СЕМЕННИК
Макроструктура
Мужские половые железы — семенники, или яички, testes — у кобелей и котов относительно небольшие. После достижения половой зрелости их размеры больше не изменяются. По форме семенники у кобелей овоидные, слегка сжатые, у котов — почти круглые. На головчатом конце, extremitas capitata, и на хвостатом конце, extremitas caudata, головка и хвост придатка прочно срастаются с семенником. Придатковый край, margo epididymalis, частью брыжейки семенника подвижно соединен с придатком, так что образуется семенниковая сумка, bursa testicularis, открытая латералыю. Обе боковые поверхности, латеральная и медиальная, facies lateralis et facies medialis, и свободный край, margo liber, покрыты брюшиной висцеральной пластинкой, lamina visceralis, влагалищной оболочки, tunica vaginalis. Под ней лежит капсула — белочная оболочка семенника, tunica albuginea testis — в виде грубоволокнистого слоя. В белочной оболочке проходят ветви семенниковой артерии, a. testicularis. У кобеля ветви семенниковой артерии извиваются лишь слегка, у кота они еще более выпрямлены (Gutzschebauch. 1936). Вместе с ветвями семенниковой вены артерия образует внутри капсулы сосудистый слой, stratum vasculare. От капсулы отходят тонкие перегородки семенника, septula testis, которые направляются к средостению семенника, mediastinum testis. Перегородки разделяют паренхиму семенника на дольки, lobuli testis, в которых расположены извитые семенные канальцы, tubuli seminiferi contorti. Последние переходят в прямые семенные канальцы, tubuli seminiferi recti, которые в свою очередь вливаются в сеть семенника, rete testis, находящуюся в средостении. От сети отходят выносящие канальцы семенника, ductuli efferentes testis, в головку придатка.
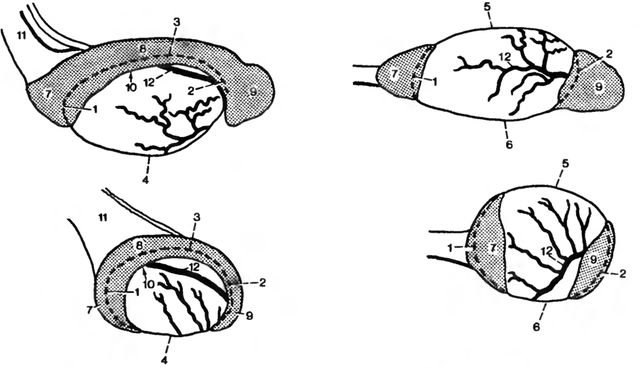
Рис. 1. Семенник и придатки семенника у кобеля (вверху) и кота (внизу). Слева: вид сбоку, справа, вид снизу (по Gutzschebauch, 1936)
1-6 testes: 1 extremitas capitata, 2 extremitas caudata,3 margo epididvmalis, 4 margo liber, 5 facies lateralis, 6 facies medialis; 7 — 9 epididymis: 7 caput epididymidis, 8 corpus epididyniidis, 9 cauda epididymidis; 10 доступ к bursa testicularis, 11 funiculus spermaticus; 12 a. testicularis и ее ответвления
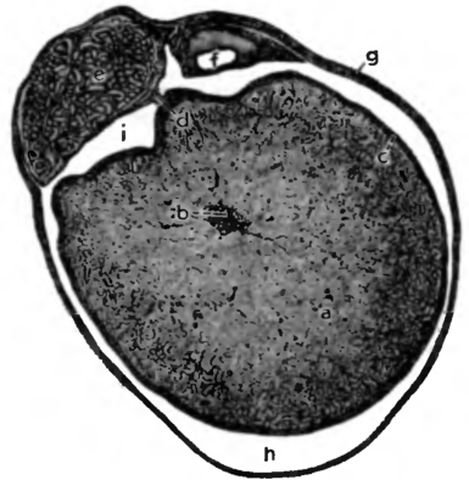
Рис. 2. Processus vaginalis с семенником, придатком семенника и семявыносящим протоком у кобеля, поперечный разрез (по Schummer, 1987)
a testes, паренхима; b mediastinum testis et rete testis; с tunica albuginea, покрытая висцеральной пластинкой влагалищной оболочки; d mesorchium; е corpus epididimidis с петлей ductus epididymidis; f ductus deferens, g processus vaginalis, состоящий из fascia spermatica interna и lamina parietalis влагалищной оболочки; h cavum vaginale, расширена; t bursa testicularis
Строение семенных канальцев
Диаметр извитых семенных канальцев у кобеля составляет 200 — 210 мкм (Foote et al., 1972), у кота — 150 —240 мкм (Ellenberger, 1911; Pier, 1985; Prem, 1992). Специальный концевой сегмент соединяет каждый извитый семенной каналец с прямым семенным канальцем.
Стенка извитого семенного канальца состоит из трех компонентов: базальной мембраны, опорных клеток, или клеток Сертоли, и гоноцитов—сперма-тогенного эпителия на разных стадиях развития.
Толщина базальной мембраны у плотоядных, как и у других домашних животных, составляет около 3, 5 — 7 мкм. За базальной мембраной спер-матогенного эпителия следует тонкий слой коллагеновых и эластических волокон.
Клетки Сертоли, или опорные клетки, представляют собой соматические компоненты эпителия канальцев. Они широким основанием лежат на базальной мембране и пронизывают весь спер-матогенный эпителий. Их многочисленные тонкие отростки охватывают клетки различных слоев сперматогенного эпителия. На поперечном срезе стенки канальца можно обнаружить от 15 до 25 равномерно распределенных опорных клеток. При окраске гистологического препарата обычными методами ядра клеток Сертоли у собаки и кошки имеют продольно-овальную или треугольную форму. Они располагаются в базальной трети сперматогенного эпителия. Ядерная мембрана имеет складки. В большинстве случаев можно распознать хорошо сформированное ядрышко.
Клетки Сертоли являются мультифункцио-нальными: они выполняют в сперматогенном эпителии питательные, защитные и координирующие функции. Кроме этого, они фагоцитируют вещества, накапливающиеся в результате сперматогенеза и расщепляют их с помощью лизосомальных энзимов. Отростки соседних клеток Сертоли соединяются друг с другом в базальной трети эпителиального слоя посредством платных контактов ("tight-junctions”) и разделяют таким образом эпителий на базальные слои, содержащую сперматого-нии и прелептогенные сперматоциты, и на адлюми-нальную (околопросветную часть), в которой происходят дальнейшие стадии сперматогенеза. Соединения между соседними клетками. Сертоли образуют барьер, благодаря которому в адлюминальной части, где происходит мейоз и дифференциация сперма-тид, создаются специфические условия. Продукты секреции клеток Сертоли образуют основную часть канальцевой жидкости. Это трансферрин, андроге-нобразующий белок, инозитол и глутамин. С этой жидкостью, помимо прочего содержащей большое количество ионов, неподвижные сперматозоиды транспортируются из семенников в придатки. Путем секреции гормона ингибина клетки Сертоли тормозят образование фолликулостимулирующего гормона в гипофизе.
Различные типы гоноцитов у собаки и кошки имеют легкие морфологические различия. Поэтому клетки, образующиеся в процессе сперматогенеза, необходимо рассматривать для обоих видов отдельно.
Гоноциты кобеля
Цикл сперматогенного эпителия разделяется на 8 (Foote et al., 1972) или 10 (Ibach et al., 1976) стадий. В качестве критерия при этом используются различия в акросомальной системе во время дифференциации сперматид и появление характерных ассоциаций гоноцитов в эпителии извитых семенных канальцев. С помощью маркировки тимидином была установлена продолжительность цикла сперматогенного эпителия — 13, 5 дней (Foote et al., 1972).
С помощью световой микроскопии у собаки можно отчетливо различать сперматогонии А и сперматогонии В (Foote et al., 1972). Сперматогонии А имеют относительно крупное овальное или круглое ядро с выраженным ядрышком. Хроматин может быть равномерно распределен, или же возможно наличие довольно крупных сгустков гетерохроматина. Сперматогонии В имеют небольшое круглое ядро. Хроматиновые агломераты прилепляются к ядерной мембране. С помощью ауторадиографических методов исследования (Ibach et al., 1976) возможно дальнейшее различение спермато-гоний собаки (A1, А2, А3, А4, Im и В).
В результате митотического деления спермато-гоний В возникают прелептотенные сперматоциты I порядка. Ядра вновь образовавшихся спермато-цитов сначала меньше ядер сперматогоний В, и их хроматин окрашивается менее интенсивно. Они располагаются ближе к базальной мембране. В стадии лептотены хромосомы начинают закручиваться в спирали и видны в ядре как нитчатые структуры. При этом ядро увеличивается, в нём возникают области, свободные от хроматина. В стадии зиго-тены гоноциты отходят дальше от базальной мембраны. Хромосомы укорачиваются. В стадии пахите-ны сперматоциты I порядка характеризуются сетчатым распределением хроматина. Размеры ядра составляют 10 мкм. В стадии диакинеза ядерная мембрана полностью разрушается: Затем проходит первое деление мейоза и гомологичные хромосомы распределяются в двух сперматоцитах II порядка, у которых, таким образом, получается гаплоидный набор хромосом. Ядра вторичных сперматоцитов (II порядка) круглые. Их диаметр равняется 7, 3 мкм. Хроматин распределен в нуклеоплазме равномерно. Вторичные сперматоциты обнаруживаются только на 4-й стадии цикла (Foote et al., 1972). Средняя продолжительность их жизни составляет полдня. Минуя фазу синтеза ДНК, вторичные сперматоциты вступают во второе деление мейоза. В результате этого получаются сперматиды с круглыми ядрами. Размеры их ядер составляют 5 мкм, что заметно меньше, чем ядра вторичных сперматоцитов. Количество ДНК в них вдвое меньше, чем в сперматоцитах II порядка, т. к. каждая хромосома состоит из одной хроматиды. Развитие круглоядерных сперматид в зрелые сперматозоиды происходит в течение 10, 5 дней (Foote et al., 1972) и включает в себя обычные фазы (фаза Гольджи, фаза колпачков, акросомная фаза и фаза созревания/зрелости).
Гоноциты кота
Подробное описание сперматогенеза можно найти у Pier (1985) и Prem (1992). Далее приведены основные характерные признаки отдельных генераций гоноцитов, наблюдаемые при помощи световой микроскопии.
Сперматогонии можно разделить на спермато-гонии А, промежуточные сперматогонии (Im-спер-матогонии) и сперматогонии В. Величина сперма-тогоний равняется 9 — 12 мкм. Сперматогонии А своей продольной осью ориентированы параллельно базальной мембране и прилегают к ней. Ядро крупное, круглой или овальной формы. Хроматин распределен гомогенно, лишь у ядерной мембраны можно раличить более выраженную кромку из гетерохроматина. Хорошо видно круглое ядрышко, расположенное в центре. В ядрах Im-сперматого-ний гетерохроматин в нуклеоплазме распределен неравномерно, в виде глыбок различной величины, имеются выраженные темные ядрышки. Под ядерной мембраной всегда располагается хорошо сформированный целостный слой гетерохроматина. Сперматогонии В представляют собой овальные клетки, которые зачастую контактируют с базальной мембраной лишь небольшим участком поверхности. Ядро круглое, содержит многочисленные грубые глыбки гетерохроматина.
Сперматоциты I порядка получаются в результате митоза из сперматогоний В. После фазы роста они переходят в состояние мейоза. Вследствие большой продолжительности профазы первого деления сперматоциты I порядка образуют большую часть всей популяции гоноцитов.
Сперматоциты стадии пролептотены обладают круглым ядром с хроматином, равномерно распределенным в виде мельчайших гранул. Нередко имеются одно-два ядрышка. В стадии лепто-тены начинается профаза первого деления созревания (I деления мейоза). Хроматин ядра выглядит более рыхлым. В стадии зиготены ядро постепенно увеличивается. Хроматин в ядре демонстрирует типичное распределение в виде полумесяца, в результате чего в остальной нуклеоплазме возникает широкая зона, свободная от хроматина. В стадии пахитены вся клетка заметно увеличивается в размерах. В ядре концентрирующийся хроматин образует грубые сетчатые структуры. В стадии диплотены ядро достигает своей максимальной величины. Хромосомы имеют вид тонких линий. Во время диакинеза происходит расхождение гомологичных хромосом. Ядерная мембрана полностью разрушается. Клетки делятся первым делением созревания (первым делением мейоза).
В результате первого деления созревания возникают сперматоциты II порядка, размеры которых составляют примерно 9 мкм и которые уже имеют гаплоидный набор хромосом. Так как они быстро переходят во второе деление созревания, их относительно редко удается обнаружить в ходе гистологического исследования. Эти клетки располагаются внутри эпителия далеко от просвета канальца. Часто в центре этих клеток имеются круглые ядрышки с плотными скоплениями хроматина.
В результате второго деления созревания (II деления мейоза) образуются круглоядерные сперматиды, которые в ходе довольно продолжительного процесса дифференциации, обозначаемого как спермиоге-нез (ранее спермиогистогенез), превращаются в сперматозоиды. Важнейшими процессами при этом являются образование акросомы, конденсация хроматина, развитие жгутика, реорганизация цитоплазмы и клеточных органелл и отделение лишней цитоплазмы. Как и у представителей других видов, у котов при этом различают фазу Гольджи, фазу колпачков, акро-сомную фазу и фазу зрелости.
Межканальцевая ткань
Межканальцевая ткань у кота большей частью состоит из клеток Лейдига. Эти большие полигональные клетки имеют круглые, частично также неправильной формы ядра, которые часто локализуются в цитоплазме эксцентрично. Цитоплазма содержит большое количество гладкой эндоплазмати-ческой сети, многочисленные митохондрии трубчатого типа и большое количество жировых капель. У кота клетки Лейдига помимо межканаль-цевой области можно встретить в белочной оболочке, в средостении семенников и в области выносящих канальцев (гетеротопные клетки Лейдига) (Mosimann/Leiser, 1985). По сравнению с котом число клеток Лейдига у собаки заметно меньше. Однако точных количественных данных для обоих видов пока нет. У собаки полигональные клетки Лейдига располагаются небольшими группами преимущественно в клинообразных пазухах между тремя соседними извитыми семенными канальцами.
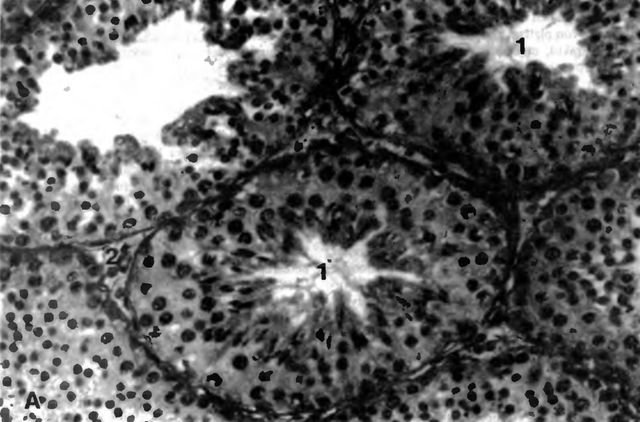
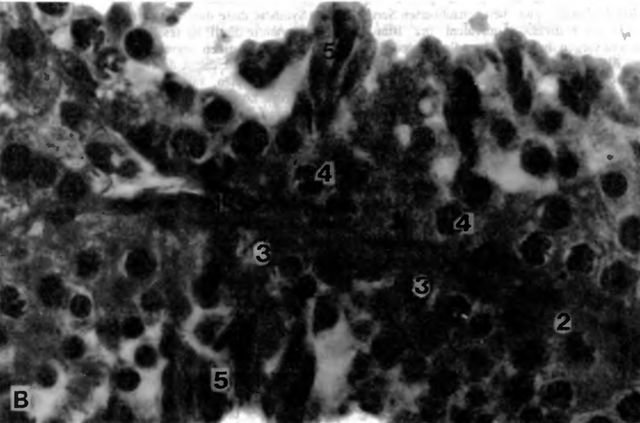
Рис. 3 А. Паренхима семенников кобеля (но Montkowski, 1993)
1 tubuli seminiferi contorti; 2 клетки Лейдига; примерно 280-кратное увеличение
Рис. 3 В. Сперматогенный эпителий кобеля (по Montkowski, 1993)
1 lamina propria; 2 клетки Сертоли; 3 сперматогонии; 4 сперматоциты I порядка; 5 поздние сперматиды; примерно 850-кратное увеличение
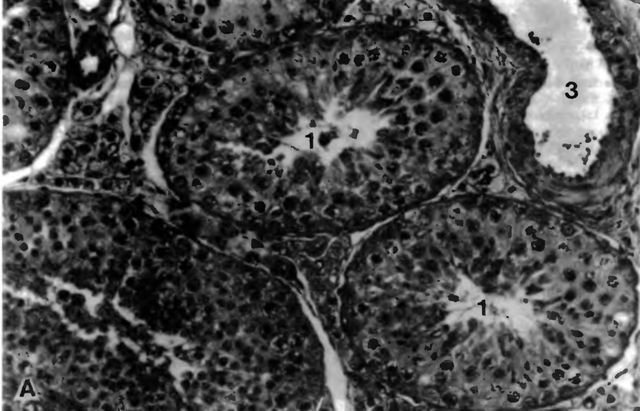
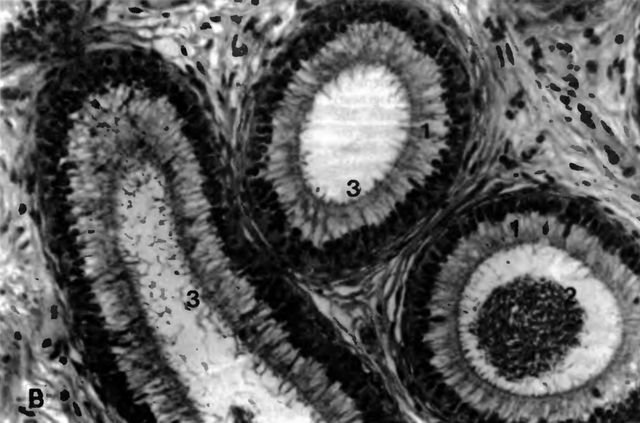
Рис. 4 А. Паренхима семенников кота
1 tubuli seminiferi contorti; 2 клетки Лейдига; 3 кровеносный сосуд, примерно 220-кратное увеличение
Рис. 4 В. Разрез через ductus epididymidis кота в области тела придатка семенника 1 ductus epididymidis, эпителий; 2 спермии, 3 стереоцилии; примерно 200-кратное увеличение
Эндокринная регуляция сперматогенеза, а также прочих функций мужской половой системы и особенностей полового поведения осуществляется преимущественно андрогенами, которые вырабатываются и накапливаются в клетках Лейдига. Выделение гормонов происходит не только васкуляр-но, то есть через кровеносные сосуды, но и через семявыводящие пути с последующим всасыванием в придатках семенников. Более подробно синтез и обмен андрогенов описывается в учебниках по физиологии (например, Bamberg, 1987).