Каждый вид существует в природе в форме своих разновидностей, а последние слагаются из популяций, свойственных отдельным биотопам, стациям. Популяция состоит из большего или меньшего числа особей. Следовательно, вид в конечном счете представляет собой совокупность особей, обладающих определенной видовой качественной спецификой, но и не равноценных между собой. Вследствие существования в несходных условиях и особи вида несколько различаются между собой как по наследственности, так и пластичности (способности дальше изменяться под влиянием внешних условий).
Существующие в природе особи испытывают на себе влияние разнообразных условий — абиотических и биотических. Возможности существования индивида более узки, чем популяции или вида, состоящих из значительного количества различающихся между собой особей. Индивид обладает избирательным потреблением элементов среды, в соответствии со своей наследственностью и способностью к адэкватной изменчивости, в пределах приспособительных возможностей. Если действующие факторы среды выходят за пределы этих возможностей, то индивид погибает, хотя его сородичи могут продолжать существовать и в меру своих индивидуальных способностей приспособительно изменяться.
Т. А. Работнов (1950), предлагающий использовать широко применяемый зоологами термин «популяция» и в ботанике («совокупность особей вида, произрастающих в определенном ценозе»), различает следующие типы популяций: I — инвазионного типа (растение находится в процессе приживания и не завершает в ценозе полного цикла своего развития; популяции состоят из занесенных извне семян, всходов или взрослых вегетативных особей); II — нормального типа (растение полностью заканчивает в ценозе спой жизненный цикл — от прорастания до образования семян; популяции состоят из семян и генеративных особей, а у многолетников— из растений па разных стадиях развития); III—регрессивного типа (генеративное возобновление растения прекратилось; популяции состоят из растений, дающих семена, подрост которых гибнет, или цветущих, но не дающих семян, или утративших способность цвести, наконец, находящихся в анабиотическом состоянии).
С. А. Северцов широко исследовал воздействие внешней среды на численность популяции. Численность популяции зависит от плодовитости и смертности. Следует различать массовую смертность, вследствие эпизоотий и стихийных бедствий (имеющих свою периодичность, различную для разных групп), от смертности индивидуальной, происходящей от незаразных болезней и от взаимодействия жертвы и хищника.
Количественное изучение индивидуальной смертности и вычисление кривых смертности во времени позволяет выяснить коэффициенты смертности, которые специфичны для каждого вида и каждой возрастной группы, так как они определяются степенью адаптации. Различие в коэффициентах смертности у разных животных объясняется различными типами размножения (яйценоскость, живородность, забота о потомстве). Между высотой типа размножения и коэффициентом смертности существует определенная корреляция: чем выше приспособление и охрана зародыша от гибели, тем ниже коэффициент смертности.
Следовательно, форма и активность борьбы за существование менялись в течение эволюции в связи с разными направлениями морфофизиологического прогресса, как это показано С. А. Северцовым в отношении рыб, рептилий и млекопитающих. Устанавливается некоторая закономерность: чем выше морфофизиологический уровень группы животных, тем ниже коэффициент гибели и меньше количество производимых детенышей (Матвеев, 1936).
Внутривидовые отношения организмов, направленные в основном на обеспечение размножения особей, весьма разнообразны и связаны с выработкой различных приспособлений, имеющих своим назначением наиболее успешное выполнение этой жизненно важной функции. Внутривидовые отношения организмов, как правило, содействуют лучшему выживанию вида, которое обеспечивается увеличением его численности.
Поскольку внутривидовые отношения организмов складываются вокруг размножения и имеют своей биологической «целью» возможное максимальное увеличение численности особей вида и его расселение, то необходимо рассмотреть различные формы размножения организмов и выяснить, какие имеются приспособления к скорейшему повышению плотности популяции. Этот материал позволяет обосновать новыми данными критику творческим дарвинизмом мальтузианских идей об особом значении перенаселенности в мире организмов.
Для животных и растений характерны две основные формы размножения— бесполое и половое, причем вторая биологически более прогрессивна. Половое размножение, в свою очередь, может происходить без оплодотворения (партеногенетически) или с таковым. Как известно, еще Ч. Дарвин открыл закон биологической полезности перекрестного оплодотворения, действие которого объясняет возникновение разнообразных приспособлений, направленных к обеспечению нормального полового процесса.
Бесполое, или вегетативное, размножение широко распространено в мире простейших, а также многоклеточных растений и животных. Осуществляется оно в основном путем деления и почкования, с отделением индивидов или образованием колоний.
Колонии, наблюдаемые, например, у губок и полипов, представляют собой совершенно своеобразные внутривидовые явления. В зависимости от характера дифференцировки членов колонии различают мономорфные (с одинаковыми особями — губки, красный коралл) и полиморфные колонии (с разделением труда между особями — гидроидный полип).
Своеобразие колоний как биологического явления заключается и стирании граней между индивидами. На это обратил внимание еще Ф. Энгельс, который писал: «У низших животных невозможно строго установить понятие индивида; не только в том смысле, является ли данное животное индивидом или колонией, но и по вопросу о том, где в процессе развития прекращается один индивид и начинается другой... ».
Правильное чередование поколений, или метагенез, характеризуется сменой бесполого размножения половым (например, у некоторых медуз). В этом случае два поколения одного вида морфологически отличаются необычайно резко (медуза и полип), живут в разных биотопах и обладают разными требованиями к условиям жизни. Этот факт говорит о том, что в качестве видового критерия возможность размножения имеет большее значение, чем простое сходство признаков. Поэтому для внутривидовых отношений характерна обеспеченность нормального размножения.
Еще более разнообразные взаимоотношения возникают между организмами на почве полового размножения. Здесь можно различать партеногенез (девственное размножение женских особей без оплодотворения), гамогенез (размножение с оплодотворением) и гетерогонию (смену партеногенеза гамогенезом).
Партеногенез наблюдается у ряда растений и животных. В некоторых случаях он является единственным способом размножения (отдельные виды орехотворок, тлей и круглых червей), но чаще чередуется с гамогенезом.
Гетерогония наблюдается у коловраток и ветвистоусых ракообразных, вызывая у них сложный годовой цикломорфоз, состоящий в смене одного, двух или нескольких двуполых поколений многими однополыми.
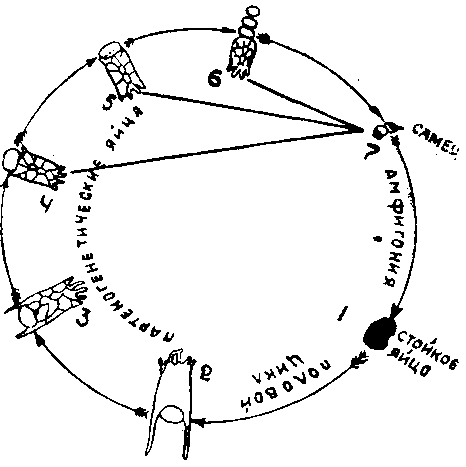
Рис. 1. Цнкломорфоз коловратки Anuraea: 1 —стойкое яйцо, 2 — A. aculeata typica, 3 — A. aculeata, 4 — A. a. brevisptina. 5 — A. a. valga, 6 — A. a. curvicornis, 7— самец (у 2—5 партеногенетические яйца, дающие самцов).
Интересно отметить, что у названных гидробионтов происходит ограничение партеногенеза на севере.
Увеличение количества партеногенетических поколений при благоприятных условиях жизни является, несомненно, полезным для вида приспособлением, обеспечивающим скорейшее размножение и заселение особями всего возможного местообитания. Обычное двуполое размножение задержало бы возрастание численности особей вида, так как половина их была бы представлена самцами и требовалось известное время на процесс оплодотворения. Факты цикломорфоза интересны как доказательство наличия в природе недонаселенности и выработки у некоторых видов специальных приспособлений для ускорения темпа размножения и прогрессивного увеличения численности популяции.
Дальнейшее приспособление в этом отношении представляет открытый Н. П. Вагнером педогенез, или детское размножение — способность личинок некоторых насекомых (мелких мушек хирономид) преждевременно достигать половой зрелости и размножаться партеногенетически.
С точки зрения своеобразия внутривидовых отношений явления партеногенеза и гетерогонии должны привлечь внимание тем, что здесь популяция длительное время (несколько месяцев в году) может быть представлена особями одного пола. В такой популяции взаимоотношения полов будут сведены к минимуму.
При размножении с оплодотворением — гамогенезе, в свою очередь, наблюдаются такие отклонения, как гермафродитизм (слитнополость, с самооплодотворением или перекрестным оплодотворением). В целях обеспечения последнего, дающего потомству большую жизненность, у гермафродитов наблюдается разновременность созревания мужских и женских половых клеток внутри организма. Тем самым исключается возможность самооплодотворения. Но законен вопрос, в чем же в таком случае смысл гермафродитизма, если природа препятствует осуществлению самооплодотворения? Среди животных явление гермафродитизма распространено преимущественно у сидячих форм и паразитов, где оно обеспечивает возможность существования вида, так как условия их жизни особые (трудность встречи самцов и самок, невозможность избежания гибели при действии неблагоприятных условий, малая выживаемость потомства). Совмещение в одной особи самца и самки сразу как бы удваивает численность популяции и является важным фактором повышения выживаемости вида. Два гермафродитных организма заменяют четыре обыкновенных (двух самцов и двух самок), и с этим нельзя не считаться.
При наличии раздельнополости ускорение размножения достигается путем неотении — созревания и оплодотворения личинок (наблюдается у жаброносного аксолотля).
При различных формах полового размножения всегда можно заметить общую тенденцию, заключающуюся в том, что имеются приспособления к возможному укорочению жизненного цикла и повышению плодовитости особей. При улучшении условий жизни организм обычно начинает быстрее расти, раньше созревает и становится плодовитее. Все это говорит о том, что виды, как правило, не страдают от перенаселенности и потому имеют разнообразные приспособления, направленные не к ограничению численности потомства, а к всемерному—насколько позволяет производительность организма — его увеличению.
На почве полового размножения у животных возникает ряд особых взаимоотношений, проявляющихся в создании временных или постоянных семейных группировок. Сюда относится образование: 1) постоянных или временных (на период размножения) «гнезд» из двух или нескольких производителей (рыба колюшка, скворцы, лебеди и т. д. ); 2) различных «стад» — стай саранчевых и птиц, косяков рыб, стай млекопитающих и т. п., которые возникают на период миграции или размножения; 3) постоянных полиморфных «общин»—у шмелей, ос, пчел, муравьев и термитов.
Все эти явления, а также и сложные инстинкты, связанные с так называемой заботой о потомстве, направлены к одному — обеспечению возможно большей выживаемости особей своего вида, так как, очевидно, в природе существует много ограничивающих факторов, помимо пищи, которые непрерывно сокращают население вида. И если бы у видов не было разнообразных приспособлений к возможно большему продуцированию яиц и лучшему выживанию потомства, то существование и эволюция видов стали бы невозможны. Таким образом, никакого антагонизма между особями одного вида в природе, как правило, не наблюдается.
У высших беспозвоночных и позвоночных животных, в. связи с более активной (физиологически) деятельностью половых органов и появлением сложной нервной системы, развиваются вторичнополовые «признаки. Наличие последних обусловливает явление так называемого полового диморфизм а (у некоторых — полиморфизма).
Вторичнополовые признаки обеспечивают лучшее размножение вида и потому имеют тенденцию к закреплению и усилению у потомства. Особая форма внутривидовых отношений, связанная с образованием вторичнополовых признаков у животных, была названа Ч. Дарвином половым отбором. Как известно, половой отбор характеризуется тем, что побежденный самец, как правило, не лишается жизни, но лишь ограничивается в своей половой функции. Следовательно, и в том случае, когда внутривидовые отношения животных приводят к конкуренции или прямой борьбе самцов из-за самок, взаимного истребления все же не происходит, Погибают лишь явно нежизнеспособные особи, существование которых не в интересах вида.
Отсутствие вторичнополовых признаков и связанного с ними полового отбора у растений и низших беспозвоночных, показывает наличие существенных различий между внутривидовыми отношениями в отдельных группах организмов. В то же время действие полового отбора усиливается с повышением организации животных в пределах отдельных групп, например, у позвоночных — от рыб к млекопитающим.
Много специфичного таят в себе и внутривидовые отношения, возникающие в некоторых случаях на почве питания, например, явление каннибализма и т. п., которые уже упоминались выше. В нашей современной зоологической литературе они трактуются как видовые приспособления к специфическим условиям существования, характеризующимся недостатком пищи. В таких случаях внутривидовые хищничество и паразитизм оказываются полезными виду, так как при их отсутствии определенные местообитания оставались бы для данного вида непригодными. Например, окунь может жить в безрыбных озерах лишь при каннибализме взрослых особей.
Специфика внутривидовых отношений на почве питания у животных, которые нельзя отнести ни к борьбе, ни к взаимопомощи, может быть проиллюстрирована на следующем наглядном примере.
В конце декабря и в январе на Иссык-куле наступает похолодание. В прибрежных участках озера образуется припай шириной до 100—150 м и больше, при толщине до 30—50 см. Восточный угол озера покрывается льдом почти сплошь на период до апреля. Постепенное нарастание ледяного покрова, естественно, отрицательно влияет на жизнь зимующих здесь птиц.
С мелководных участков, богатых пищей, притом легко добываемой птицами, они вытесняются льдом в менее благоприятные участки— в открытую, более глубоководную часть озера, не защищенную от постоянно дующих ветров. В результате этого происходят перекочевки части птиц в другие районы, где за счет этих переселенцев временно увеличивается плотность птичьего населения и начинает сказываться недостаток в пище.
Это случайное, зависящее от климатических условий, перенаселение птиц влечет за собой ослабление их жизнедеятельности. У птиц происходит постепенное снижение их упитанности, достигнутой в первый период зимовки, а некоторые особи со временем истощаются настолько, что они или не в состоянии улететь в район гнездования, или, улетев туда, не гнездятся. В местах временного перенаселения во второй половине зимы наиболее выносливым (крепким), упитанным) птицам приходится добывать корм с все больших глубин (5—6 м). Слабые особи красноносого нырка (больные, пораженные глистами и пр.), не способные добывать пищу с таких глубин, продолжают еще некоторое время держаться в этих стациях, прибегая к особым уловкам.
Птица, способная добыть корм с больших глубин, ныряет за ним, всплывая затем на поверхность воды с лучком хары в клюве. К ней тотчас бросаются 3—5 ослабевших особей, которые моментально растаскивают добытый корм, причем добытчику остаются лишь обрывки водорослей, находившихся непосредственно в клюве. Такой способ питания не может полностью удовлетворить потребность птиц в пище, и от систематического недоедания происходит постепенное ослабление организма, что отражается на снижении упитанности птиц.
По поводу описанных внутривидовых взаимоотношений, наблюдающихся у красноносого нырка в условиях временной перенаселенности в наиболее тяжелый период года, следует сказать, что они действительно не могут быть отнесены ни к борьбе, ни к взаимопомощи. Очевидно, что при «борьбе» должен быть «победитель» и «побежденный», а при «взаимопомощи» должна быть обоюдная польза, как это наблюдается при взаимоотношениях особей разных видов. Но здесь нет ни того, ни другого, а выступает налицо качественное своеобразие внутривидовых пищевых взаимоотношений, заключающееся в общем ослаблении организмов в связи с временной перенаселенностью, причем происходит полезное для вида выживание наибольшего количества особей, с естественным отпадом наиболее ослабленных. При этом гибель ослабленных вызвана прямым и косвенным действием отрицательных абиотических условий и межвидовых отношений.
Так называемая внутривидовая конкуренция здесь ни при чем. Пищи достаточно, но временно ограничена, затруднена возможность ее доставания и это создает перенаселенность, в результате которой происходит гибель части особей — ослабленных. Гибель — следствие прямого влияния неблагоприятных условий, но не конкуренции, совместное же обитание спасает даже слабых, которые одни не могли бы пропитаться.
На одиночную особь действуют различные абиотические и биотические факторы, многие из которых легко могут оборвать ее индивидуальную жизнь. Дарвин указывал, что редкость формы является предвестником ее вымирания.
Абиотические условия в большинстве случаев одинаково действуют как на одиночную особь, так и на популяцию. Например, низкая или высокая температура, недостаток или избыток влажности, кислорода, солености, света и т. п. будут одинаково влиять как на индивид, так и на группу индивидов. В некоторых случаях, когда речь идет о потреблении абиотических факторов, например, кислорода, воды и т. п., в популяции недостаток наступит лишь скорее, но не изменится направление процесса, как указывал еще Ф.Энгельс.
Иначе действуют на индивид и популяцию биотические условия. Одиночная особь не может долго существовать в условиях межвидовой борьбы за существование, популяция легче противостоит ей. Там, где имеет место большая гибель особей на разных стадиях индивидуального развития, популяция сохраняется благодаря наличию приспособлений к повышению плодовитости и выработке ряда других особенностей.
Популяция как совокупность особей вида в определенном местообитании противостоит окружающим неблагоприятным условиям, как абиотическим, так и биотическим межвидовым. В целом ряде случаев особям вида выгодна групповая жизнь, /постоянно или временно, и это ведет к образованию соответствующих популяций.
У прикрепленных растений и подвижных животных в этом отношении есть и сходство, и различие. Те и другие могут выжить лишь при огромном изобилии зародышей (этим объясняется высокая плодовитость, в норме обеспечивающая выживание лишь исходного количества производителей). Гнездовой посев и групповой выпуск животных являются важнейшими условиями закрепления организмов в новом местообитании. Таково сходство. Различие же между растениями и животными состоит в следующем.
Растения, прикрепленные к месту, по мере роста и увеличения размеров создают перенаселенность (или видимость ее), и ботаники усматривают в ней лишь конкуренцию, не замечая пользы от известной численности популяции, без которой вид часто не мог бы существовать. Что же касается подвижных животных, которые могут активно избегать перенаселенности, но часто именно активно образуют густые группировки, то здесь польза повышенной численности популяции говорит сама за себя. Поэтому зоологам почти не приходится прибегать к понятию внутривидовой конкуренции, но, напротив, выявлять конкретную пользу повышенной численности внутривидовых объединений, которые обеспечивают наиболее успешное размножение, существование и расселение организмов.
В этом отношении особый интерес представляют явления временной и постоянной стадности у животных. Образование стай, колоний, кулиг, стад и т. д. свойственно разным видам и, несомненно, полезно для выживания вида.
А. Г. Скопцов (1953) показал, что в группе личинок насекомых, ведущих объединенный образ жизни (златогузки, большая пчелиная моль), температура всегда выше, чем в окружающей среде. Выделение тепла гусеницами при движении создает в гнезде особый микроклимат. В группе гусеницы растут быстрее, чем при одиночном воспитании. Измерения интенсивности дыхания показали, что уровень обмена в группе всегда ниже, чем у одиночных особей. Таким образом, групповой образ жизни для этих личинок благоприятен и он наследственно закрепился как полезное приспособление.
В. В. Шулейкин (1941) установил, что стая птиц в полете, как и косяк рыб в движении, представляют закономерную механическую систему. Мелкие птицы образуют в полете воробьиную стаю, крупные — журавлиный клин. Во время движения такой стаи все особи в ней связаны пондеромоторными силами и не могут от нее отбиться — их притягивает к стае. Движущийся косяк рыб состоит из особей одного вида и размера, обладающих одинаковой мышечной силой разношерстное скопление не могло бы двигаться монолитной массой.
Стайное поведение, как указывает Г. В. Никольский (1953), представляет видовое приспособительное свойство животных. Лучше всего изучено оно к настоящему времени у рыб, в связи с практической потребностью в соответствующих знаниях для рыболовства.
Представление о том, что стая есть результат одностороннего воздействия того или иного фактора среды, которые «сгоняют» и «отсортировывают» рыб по размерам, безусловно, ошибочно и должно быть оставлено. Биологическое значение стаи не остается постоянным для данного вида, оно может меняться как в процессе онтогенеза, так и в течение года. В связи с изменением биологического значения стаи меняется как строение, так и поведение особи.
В связи с полезностью стайного образа жизни у рыб имеются морфологические приспособления к нему, заключающиеся в наличии стайной окраски (ярких полос и пятен), которые облегчают рыбам ориентироваться друг на друга. С возрастными изменениями стайности находятся в связи возрастные изменения окраски рыб, например, молодь амурского горчака стайна и имеет черное пятно на спинном плавнике, которое позднее исчезает; в Амуре кета в период нереста получает темную полосу на хвосте.
Биологическое значение стайности у рыб разнообразно. У ряда видов рыб, как и у других животных, стая имеет защитное значение. В стае рыбы на более далеком расстоянии обнаруживают опасность и во многих случаях стаей лучше защищаются от врагов. Если в стаю рыб капнуть сок тела или бросить раздавленную особь данного вида, то стая рассредотачивается (защита от хищника). Стаи рыб с более далекого расстояния замечают орудия лова и легче их избегают. Уловы активных орудий лова часто выше при добыче разреженных рыб. Стая легче уходит из ловушки.
Стая имеет значение у многих видов для отыскания кормовых объектов. В стае те рыбы, у которых стайный образ жизни имеет значение для питания, питаются обычно с большей интенсивностью, чем в одиночку. Рыбы, питающиеся в пелагиали планктоном и рыбой, в стае питаются интенсивнее. Пищи в окружающей среде много, и это биологически выгодно для вида. Скорость прохождения пищи через кишечник у стайной и одиночной рыбы также различна. У рыб, питающихся бентосом, более активное питание в одиночку ведет к сохранению их кормовой базы.

Скопление бабочек боярышниц (Aporia crataegi), пьющих воду на увлажненной почве. Долина р. Калтат, Красноярский край. 22 июня 1954 г.

Поселение SemiculcDspira libertina и Balanus sp. на литорали острова Монерон
Японское море. Август 1957 г,

Личинки ручейников (Trichoptera) на зимовке. Верховья р. Маны. Ноябрь 1953 г.

Лежбище сивучей (Eumetop'ias jubatus) на Курильских островах 12 сентября 1956 г.
Несомненно, имеет место миграционное значение стаи. Стая легче ориентируется и находит миграционный путь. Зимовальное значение стаи выражается в обеспечении, в первую очередь, более благоприятных условий обмена, в частности путем созданий «слизевой защиты». Воздействие токсических веществ на стаю ослабевает.
Изучение биологического значения стаи необходимо для того, чтобы прогнозировать поведение, рыб и находить пути управления стаей.
В ихтиологии разработан вопрос о типах нерестовых популяций у рыб (Г. Н. Монастырский), характеризующихся определенным соотношением возраста половой зрелости, плодовитости и других признаков физиологии развития. Изучаются элементарные популяции рыб (Н. В. Лебедев), представляющие собой пространственно обособленные группы физиологически однородных особей.
Весьма характерно стайное поведение у птиц. В качестве примера укажем поведение лысухи на озере Иссык-Куль, изучавшееся Ф. Ф. Пятковым.
В стае птицы оказываются и лучше защищенными от своих врагов (хотя это положение не является универсальным и не применимо ко всем случаям, так как у других видов при появлении опасности может быть выгодным рассредоточение). При тихой погоде лысухи держатся на озере небольшими группами и кормятся. Но вот появляется орлан. Заметив его, лысухи бросают кормиться, проявляют беспокойство и начинают сплываться в кучу. Птицы, находящиеся вдали, поднимаются и, не отрываясь от воды, бегут по ней, махая крыльями, спеша присоединиться к стае. Вскоре образуется плотная масса лысух, которые при приближении орлана с шумом и криком перевертываются на спину, выбрасывают ногами массу брызг, образующих сплошную водную завесу, через которую хищнику трудно увидеть и схватить какую-нибудь птицу. Как правило, жертвами орлана являются одиночные лысухи, не успевшие присоединиться к стае. Таким образом, стая птиц имеет и несомненное защитное значение.
Очевидно, что дальнейшее изучение стадности животных позволит открыть новые закономерности формирования популяций и характерных для них внутривидовых отношений. Как нами уже раньше указывалось, последние, как правило, не носят антагонистического характера. Здесь следует добавить, что у животных имеется целый ряд приспособлений, обеспечивающих возможное ослабление напряженности внутривидовых пищевых отношений: возрастные различия в пище, разница в питании самцов и самок, впадение животных в спячку и т. п. Как указывает Г. В. Никольский (1949), в сложившейся фауне рыб наименее обеспеченными пищей оказываются особи старших возрастных групп. Следовательно, причиной большой смертности на ранних этапах развития служит не недостаток пищи, а факторы абиотической среды и хищники.
Следует сказать, что, к сожалению, экология популяций животных и растений изучена еще совершенно недостаточно. Зоологи изучают преимущественно жизнь индивидов, а ботаники пытаются сразу познать биоценозы.
Популяции особей вида в разных участках ареала — или, точнее, в разных местообитаниях — неоднородны. В настоящее время можно со всей определенностью говорить о внутривидовой биологической разнокачественности популяций.
Наличие внутривидовых подразделений — упомянутых выше географических и негеографических форм, устанавливаемых на основании морфологических признаков, — свидетельствует о значительной биологической неоднородности особей вида. Этот вопрос, однако, еще слабо разработан, и для характеристики относящихся к нему материалов используем прежде всего данные современной ихтиологии.
Биологическая неоднородность вида у рыб усиливается специфическим своеобразием их среды жизни и характером местообитаний — большей и меньшей изоляцией озерно-речных бассейнов, а особенно ограниченностью нерестовых площадей многих видов.
Крупным открытием явилось установление Л. С. Бергом (1934) наличия яровых и озимых рас у проходных рыб. Не имея особых морфологических различий, помимо величины, эти расы значительно отличаются своей экологией сроками миграции, состоянием зрелости половых продуктов, районом преднерестовой зимовки, расположением нерестилищ и т. д. Яровыми расами были названы такие, которые мечут икру в том же году, когда началась нерестовая миграция, озимыми — мечущие на следующий год и зимующие в реке (растянутая нерестовая миграция с перерывом на зимовку). С этим связаны и другие отличия: у первых половые продукты во время входа в реки более зрелые, чем у вторых и т. п.
На биологическую неоднородность видов рыб издавна указывала практика рыбного промысла, различающая отдельные «стада», па нее обращали внимание и некоторые исследователи. Н. Л. Гербильский (1950) на примере, осетровых рыб развивает учение о внутривидовых биологических группах в пределах отдельного бассейна и всего ареала. «Биологической группой, —говорит Н. Л. Гербильский, — мы называем совокупность особей, относящихся к одному виду, но отличающихся от других биологических групп того же вида особенностями биологии развития. В мире рыб эти различия в первую очередь относятся к явлениям, связанным с процессом размножения (различия в половом цикле, в состоянии производителей в период их захода из морей в реки, в сроках и местах нереста, в нерестных температурах и т. п. ).
Н, Л. Гербильский иллюстрирует приложимость учения о внутривидовых биологических группах к практике рыбоводных работ на примере куринского осетра, у которого выделяются четыре группы: ранний яровой осетр, поздний яровой осетр, озимый осетр летнего хода и яровой осетр осеннего хода. Автор справедливо указывает, что обнаружение и детальная рыбоводнобиологическая характеристика биологических групп проходных и полупроходных рыб является необходимой предпосылкой дальнейшего развития их заводского воспроизводства и их использования для акклиматизационных целей.
Как показывают исследования, внутривидовые биологические группы имеются не только у проходных рыб, но и у туводных. Поэтому мы приходим к выводу, что представление о внутривидовой биологической разнокачественности касается рыб любых экологических групп — речных, озерных, морских и др. Очевидно, также, что это свойства характерно не только для рыб, но и других организмов.
И. В. Кожанчиков (1948) рассматривает вопрос о биологических формах в энтомологии. Среди многих растительноядных и хищных насекомых имеются специфические внутривидовые формы. У жуков-листоедов и короедов особи одних и тех же видов обнаруживают трудности скрещивания, территориальную изоляцию и, иногда, небольшие цветовые отличия. Биологические фомы были специально оценены и описаны как самостоятельные виды Н. А. Холодковским для хермесов и тлей, специализированных на питании определенными видами хвойных растений. Близкая к хермесам виноградная филлоксера обнаруживает сложный комплекс кормовых форм и среди них две наиболее дифференцированные и отчасти разобщенные географически. Явление биологической дифференциации у тлей выражено очень резко, и многие виды их значительно более определенно характеризуются типом повреждения кормовых растений (галлами) и их видовыми составом, чем структурами. Различающиеся своей пищей биологические формы обнаружены среди видов разных отрядов насекомых.
Наряду с пищевыми формами у насекомых известны и другие дифференцированные в отношении термического режима, по поведению или по физиологическим особенностям. Обыкновенный малярийный комap дает сложный комплекс биологических форм, отчасти разобщенных географически, но неясных по структурным отличиям. У этих форм заметны отличия по характеру яйцекладки, по окраске яиц и по отношению к животным, кровь которых они высасывают.
Из членистоногих, кроме насекомых, биологические формы описаны для клещей и ракообразных.
Биологические формы известны также среди некоторых видов высших позвоночных. Для птиц описываются биологические формы, отличающиеся по пению, пока слабо изученные. Н. И. Калабухов (1935) нашел их у лесной мыши. Равнинные и горные популяции этого вида существенно отличаются по количеству гемоглобина в крови и по числу эритроцитов.
В биологической разнокачественности групп внутри видов заложены возможности прогрессивной эволюции последних.
Настало время перекинуть мюст от систематических внутривидовых категорий к биологическим. К сожалению, систематики, изучающие морфолого-анатомические признаки внутривидовых групп и описывающие различные разновидности (подвиды, племена, расы и т. п. ), мало внимания уделяют выявлению их экологических особенностей, а рыбоводы, энтомологи, паразитологи и другие специалисты, характеризующие экологические отличия, не интересуются морфологическими признаками. Между тем нельзя себе представить, чтобы морфолого-анатомические признаки изменялись (при образовании разновидностей) без существенных изменений физиологии и экологии организмов, точно так же, как и изменения в их экологии (при образовании биологических форм) не могут не вызвать соответствующих морфологических изменений. Как известно, форма и функция взаимосвязаны, орган таков, каково его отправление.
Поэтому особи вида, приуроченные к несколько особым условиям существования (что усиливается при изоляции водоема, специфичности нерестилищ или кормового субстрата и т. п. ), изменяются прежде всего экологически и физиологически, а затем, на этой основе, у них появляются и анатомо-морфологические отличия.
Наличие внутривидовой биологической неоднородности содействует путем перекрестного оплодотворения повышению жизненности и улучшению породных качеств. Хотя бы процент таких скрещиваний в пределах популяции был и небольшим, польза от него будет значительная (как и в работах по принудительному внутри-сортовому скрещиванию растений-самоопылителей). С другой стороны, это явление устраняет возможность перенаселения и содействует возрастанию общей численности особей вида.
Дальнейшая дифференциация биологических форм приводит к столь значительным физиологическим отличиям, что они утрачивают способность к скрещиванию или дгют мало жизненных и бесплодных гибридов (ряд насекомых). Таким образом, очевидно, могут возникать новые виды, признаваемые за вполне самостоятельные в энтомологии, паразитологии и фитопатологии.
Головная вошь (Pediculus capitis), имеющая у разных человеческих рас соответствующие цветовые разновидности: (Холодковский, 1909), довольно заметно отличается от платяной вши (Pediculus vestimenti). Однако экспериментально доказано, что головная вошь путем изменения условий существования может быть превращена в платяную, что рассматривается в качестве доказательства необходимости считать их лишь биологическими формами вида человеческой вши (Кожанчиков). Жизнь головной вши в течение ряда поколений на открытой коже. ведет к формированию популяций, по экологии и структурам особей, не отличимых от обычной платяной вши. Возникает новый характер яйцекладки и прикрепления яйца, иные плодовитость, окраска и форма тела особей. Мы полагаем, что многократно превращающиеся одна в другую формы человеческой вши именно вследствие обратимости процесса являются одним видом. Не таковы ли по своей природе и некоторые превращения среди растений, которые иногда рассматриваются в качестве примеров скачкообразного видообразования?
Нам представляется, что биологические формы, возникая внутри вида как выгодное приспособление, содействующее увеличению общей численности особей вида, наряду с другими внутривидовыми формами могут приводить к образованию новых видов. Обычно требуется несколько поколений для превращения одной биологической формы в другую. Перемена пищи, изменение типа обмена веществ — основное условие образования новых видов.
Вопрос о биологических формах имеет важное практическое значение в разных отраслях народного хозяйства в связи с необходимостью применения для: борьбы с вредными и разведения полезных форм специфических средств.
Используемая литература: Основы Экологии: Учеб. лит-ра./Б. Г. Иоганзен
Под. ред.: А. В. Коваленок,-
Т.: Типография № 1,-58 г.
Скачать реферат:
Пароль на архив: privetstudent.com