Современные тенденции в изучении физиологических аспектов фитопатологии подчеркивают особое значение химической природы расовой специфичности и роли фитоалексинов с низкой молекулярной массой в устойчивости к заболеваниям. Исследования в этих областях внесли существенный вклад в познание нами взаимодействия растения и патогена. Тем не менее работы с фитоалексинами породили некоторые противоречия относительно расовой специфичности, а поиски решения обеих проблем заставляют предполагать, что пригодными, если не равнозначными, могут быть и другие определяющие факторы.
Не существует приемлемых доказательств того, что специфическая и неспецифическая расовая устойчивость определяется высокоспецифичными механизмами, уникальными в их роли супрессоров развития патогенов. Результаты опытов с фитоалексинами поддерживают тот факт, что различия между восприимчивыми и устойчивыми растениями определяются скорее скоростью и размерами ответной реакции, чем специфичностью такой реакции или антимикробной специфичностью фитоалексинов. Таким образом, расовая специфичность должна определяться генами и взаимодействием генов-регуляторов или процессами, которые регулируют ответ «когда» и «сколько».
Кроме того, аргументы относительно роли фитоалексинов в устойчивости к бактериальным заболеваниям неубедительны, а в отношении вирусных болезней фактически не существуют. Однако в случае как бактериальных, так и вирусных заболеваний существует много примеров высокоспецифичных реакций сортов на патогены и резких различий между восприимчивостью и устойчивостью. Очевидно также, что растения обладают нерасоспецифичной устойчивостью, которая характеризуется высокой эффективностью. Различные ткани одного и того же растения, одинаковые ткани на разных стадиях развития или роста могут быть иммунными, устойчивыми либо восприимчивыми к заболеванию.
Появление новых рас патогенов, устойчивых к системным фунгицидам, показывает, что механизм устойчивости, определяемый единственным компонентом (фитоалексин), должен быть в высшей степени нестабильным. В таком случае, каким образом выживают растения? Фитоалексины могли бы играть ведущую роль в устойчивости к заболеванию при условии, что они обладают действием на многочисленные звенья метаболизма, если их образуется много и каждый имеет свой сайт действия или если точка приложения их действия настолько важна, что у патогена не могут возникать мутации, изменяющие ее (подобным действием обладают защитные фунгициды). Однако аргументов в пользу этих возможностей нет.
Нестабильность системных пестицидов, определяющаяся эффективностью их действия в одном звене метаболизма, также делает проблематичным использование специфичных токсинов для поиска устойчивости. В этом подходе заключены две опасности. Первая состоит в том, что отбор даст растения с устойчивостью, зависящей от единственного сайта действия, и, следовательно, она окажется нестабильной. Вторая заключается в том, что можно упустить неспецифическую (мультигенную) устойчивость, обладающую наибольшим потенциалом стабильности.
В этой главе будут представлены доказательства возможности системной защиты растений от грибных, бактериальных и вирусных растений с помощью контролируемого заражения грибами, бактериями или вирусами. Это, конечно, не свидетельствует о высокой специфичности. Я также попытаюсь доказать, что растение, сенсибилизированное к присутствию грибного, бактериального или вирусного патогена, активирует свой потенциал множественных механизмов устойчивости к заболеванию, действующих в присутствии патогена. Что же тогда регулируют специфические гены устойчивости?
Явление
Сообщения об иммунизации растений против болезни с помощью заражения вызывающим ее патогеном, непатогенными для сорта расами возбудителей, патогенами других видов и, реже, метаболитами возбудителей имеют по меньшей мере 100-летнюю историю. Однако совсем недавно обоснованность иммунизации растений была установлена экспериментально, а с ее установлением началось прогрессирующее изучение молекулярных механизмов, ответственных за это явление, и возможностей применения его в борьбе с болезнями. Ярвуд, Круикшанк и Мэндрайк и Мюллер установили возможность иммунизации растений против заболеваний, вызываемых грибами. Затем Росс, используя вирусы в качестве индуцирующего фактора, продемонстрировал действенность иммунизации против вирусных болезней табака, фасоли и вигны. Эти работы постепенно расширились, охватывая многих хозяев, а также вирусы, бактерии, грибы, метаболиты патогенов и химические соединения. Несмотря на то что Хехт и Бэйтмэн и Мэндрайк сообщили о неспецифической устойчивости к патогенам, индуцируемой у табака грибами и вирусами, все же широкому спектру эффективности иммунизации растений, ее метаболической и генетической значимости ранние исследователи не уделяли внимания. В своих недавних сообщениях Куч и его коллеги показали, что тыквенные культуры можно иммунизировать против вирусных, бактериальных и грибных заболеваний путем заражения вирусами, бактериями или грибами. Контролируемое заражение грибом Colletotrichum lagenarium или вирусом некроза табака защищало огурец, арбуз и мускатную дыню от заболеваний, вызываемых самыми разнообразными патогенами, включая облигатные и факультативные грибы, вирусы, вызывающие локальные некрозы и системные заражения, грибы и бактерии, вызывающие увядания, пятнистости и неограниченные поражения листьев и плодов. Иммунизация эффективна против как листовых, так и корневых патогенов. У тыквенных она носит системный характер и для проявления требует лаг-периода между индукцией и проявлением. Это действительно и для феномена, отмеченного Россом в опытах с табаком и вирусом табачной мозаики (ВТМ). С иммунизацией связано появление макро-и микроскопических некрозов, вызываемых возбудителем, однако не все типы некрозов приводят к иммунизации. Механические повреждения, повреждения, вызванные химическими веществами, сухим льдом или грибными и растительными экстрактами, не дают системной иммунизации тыквенных. Заражение грибом С. lagenarium предохраняет огурец от реакции сверхчувствительности, вызываемой Phytophthora infestans — непатогенным для этого растения грибом. Однако же инокуляция P. infestans не обеспечивает защиты огурца от поражения С. lagenarium.

1. Биологический спектр эффективности системной устойчивости, индуцированной заражением листьев огурца С. lagenarium или ВНТ
Удаление первого настоящего листа (лист-индуктор) через 72—96 ч после заражения грибами или бактериями и через 48 ч после инокуляции вирусом некроза табака (ВНТ) не ослабляет иммунизации листьев, расположенных выше места ее индуцирования. Точно так же удаление листьев, расположенных выше листа-индуктора, после приблизительно равного лаг-периода не ослабляет иммунизацию удаленных листьев. Если с иммунизированного растения удалить листья, а также пазушную почку, расположенную в месте соединения черешка и стебля, то листья могут дать корешки, и растеньица будут иммунизированы.
Сигнал иммунизации в тыквенных передается при прививке и не обладает сортовой, родовой или видовой специфичностью. Эксперименты с прививкой позволяют предположить, что в точке индукции образуется какой-то химический сигнал, который передается в другие ткани и обусловливает их устойчивость. По данным Гуэдеса с соавт., для листовых патогенов эффект иммунизации выражен сильнее над листом-индуктором, чем под ним. Тем не менее, заражая листья, можно иммунизировать корни. Кольцевание черешка листа-индуктора предупреждало иммунизацию расположенных выше и ниже него частей растения. Кольцевание черешков листьев, используемых в дальнейшем для вторичного заражения, при интактном черешке листа-индуктора предупреждало иммунизацию только в листьях для вторичного заражения. Кольцевание почти полностью прекращает движение 14С-сахарозы из листа-индуктора, позволяя предполагать, что его влияние на иммунизацию определяется нарушением передвижения веществ во флоэме. Эти данные вместе с результатами экспериментов по передаче иммунизирующего фактора через прививку и сохранению иммунизации после удаления листа-индуктора служат четким доказательством того, что индукция — это результат сигнала, передаваемого из листа-индуктора. Однако постоянная подача сигнала не обязательна, и ткани, однажды получившие его, не требуют листа-индуктора для иммунизации.
Заражение первого настоящего листа огурца грибом С. lagenarium или ВНТ и последующая (через 2—3 недели) повторная инокуляция этими микроорганизмами иммунизирует растения огурца против заболеваний, вызываемых С. lagenarium, Cladosporium cucumerinum и Pseudomonas lachrymans в течение периода плодоношения (Куч и др., неопубл. данные). Иммунизирующее действие одной индукции продолжается в течение 4—6 недель, но без повторного заражения системная устойчивость теряется начиная с наиболее удаленных частей растения и все ближе к листу-индуктору. Однако растения огурца невозможно иммунизировать, если они вступили в фазу цветения или плодоношения. Устранение цветочных почек до их раскрытия не продлевает период, в течение которого возможна иммунизация (Куч и др., неопубл. данные). Это заставляет предполагать, что «биологические часы» растения, запрограммированные на репродукцию, выключают способность к иммунизации, но не устраняют ее из уже иммунизированных тканей. Таково дополнительное доказательство, что индукция иммунитета и сдерживание патогенов представляют собой разные метаболические процессы.
Концентрация инокулюма, использованного для индукции, и количество некрозов, образовавшихся на листе-индукторе, прямо связаны с размерами иммунизации до тех пор, пока не будет достигнута точка их насыщения. Единственный некроз, вызванный С. lagenarium, и небольшое число некрозов (не более 8), вызванных ВНТ на листе-индукторе, иммунизируют ткани выше точки индукции. При высоких концентрациях вторичного инокулюма иммунизация, индуцированная низким уровнем инокуляционной нагрузки, будет до определенной степени ослабевать; например, часто происходит уменьшение не числа некрозов, а их размеров. Иногда высокие концентрации инокулюма С. lagenarium вызывают образование системных симптомов на растениях. Это явление не наблюдали на иммунизированных растениях. Сокращение размеров системного заболевания при иммунизации через листья также доказано на примере растений, повторно зараженных вирусом огуречной мозаики или Erwinia tracheiphila (Бергстром и Джонсон, неопубл. данные).
Механизмы сдерживания
Иммунизация через листья эффективна против С. lagenarium или С. cucumeritium, нанесенных на поверхность листьев или инфильтрованных в них. Она ослабляет размножение P. lachrymans и инфекционную способность ВНТ, эффективна против увядания, вызываемого корневым патогеном Fusarium oxysporum f. sp. cucumerinum и листовым патогеном Е. tracheiphila (Бергстром, неопубл. данные). Представляется невероятным, что все перечисленные явления можно объяснить только одним механизмом сдерживания.
Проникание аппрессориев С. lagenarium в ткани иммунизированного растения огурца заметно ослабевает, тогда как прорастание конидий не нарушается. В то же время прорастание конидий и проникание С. cucumerinum в иммунизированные ткани растений огурца остается без изменений. С. cucumerinum внедряется в течение 18 ч, а С. lagenarium, развивающийся из толстостенных аппрессориев, требует для этого 40—60 ч. Гистологические и химические исследования позволяют предположить, что механизмы сдерживания С. cucumerinum в иммунизированных тканях имеют общие признаки с механизмами в тканях устойчивых неиммунизированных растений. Однако в случае С. lagenarium иммунизация, видимо, активирует по крайней мере еще один дополнительный механизм сдерживания в иммунизированных растениях по сравнению с неиммунизированными устойчивыми. Проникание осуществляется одинаково легко в устойчивых и восприимчивых растениях, но только у первых ассоциируется с реакцией типа сверхчувствительности. Иммунизация устойчивого растения ограничивает прони-кание и усиливает устойчивость, но не является причиной активизации сверхчувствительных реакций вокруг места проникания у восприимчивых растений.
Иммунизация сопровождается по меньшей мере трехкратным увеличением активности пероксидазы. Оно носит системный характер и отмечено у иммунизированных, не подверженных вторичному заражению тканей, удаленных от места индуктивной инокуляции. Поскольку индукция с помощью С. lagenarium, С. сuсumerinum, P. lachrymans или ВНТ вызывает это увеличение, то оно, видимо, связано с пероксидазной активностью хозяина. При иммунизации единственное пятно некроза на листе-индукторе приводит к статистически значимому системному приросту пероксидазной активности, который коррелирует с заметным усилением активности отдельных изоферментов. Повторное заражение С. lagenarium или ВНТ заметно повышает степень иммунизации и системное увеличение пероксидазы. Активность пероксидазы после контрольного заражения С. cucumerinum и С. lagenarium возрастает скорее в иммунизированных, чем в неиммунизированных растениях, и это также связано с усилением активности некоторых изоферментов. Однако со временем общая пероксидазная активность в повторно зараженных тканях неиммунизированных растений может превысить таковую в иммунизированных. Повреждение листа сухим льдом или карборундом приводит к увеличению пероксидазной активности в поврежденном листе, но оно не имеет системного характера. В то же время возрастание активности пероксидазы может быть только одним из проявлений механизма сдерживания. В неиммунизированных тканях активность пероксидазы часто выше, чем в иммунизированных тканях после вторичного заражения. Возможно, реакция наступает слишком поздно и вначале она слишком слаба.
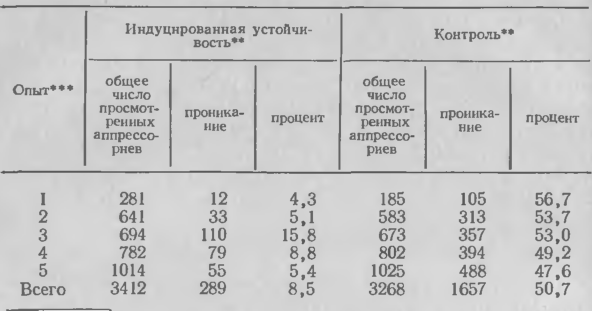
2. Влияние инокуляции первого настоящего листа сорта огурца SMR 58 грибом С. lagenarium на проникание гриба в полоски эпидермиса, вырезанные из черешков второго листа
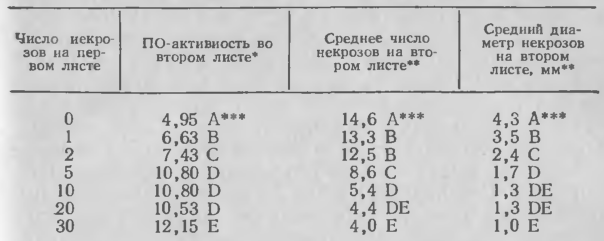
3. Влияние числа некрозов, вызванных С. lagenarium на первом настоящем листе сорта огурца SMR 58, на пероксидазную активность и защиту второго настоящего листа
Резкое усиление пероксидазной активности отмечено также в стареющих листьях огурца, причем такая ткань у неиммунизированных растений не более устойчива к С. lagenarium, чем молодые ткани. Точно так же обработка растений огурца этиленом приводит к увеличению пероксидазной активности, но не вызывает системного иммунитета. Можно дать по меньшей мере два объяснения этим явлениям. Одно заключается в том что иммунизация не требует увеличения пероксидазной активности. Это толкование совпадает с недавними сообщениями Надольны и Секейра, описавших результаты исследований по иммунизации табака. Однако, если рассматривать увеличение активности пероксидазы in vivo, большое значение имеют концентрации и доступность двух субстратов пероксидазы — перекиси водорода и соответствующего донора водорода, обычно фенола или амина. Первое объяснение и сообщение Надольны и Секейра было бы правильным, если бы пероксидаза сама по себе была бы токсичной для возбудителя или служила единственным ограничивающим его фактором. Второе объяснение состоит в том, что пе-роксидаза — это один из компонентов механизма сдерживания патогенов в иммунизированных растениях. При иммунизации усиление пероксидазной активности должно сопровождаться увеличенным или неограниченным снабжением перекисью водорода и донором водорода. Но ни стареющие листья, ни листья, обработанные этиленом, в которых не возникает иммунитет, не могут обеспечить достаточного количества этих необходимых компонентов. Кроме того, локализация и время проявления пероксидазной активности, соотнесенные с развитием патогена, могут иметь важное значение. В иммунизированных тканях активность фенилаланинаммиаклиазы (ФАЛ), р-кумарил-КоА-лиазы или р-кумарил-ре-дуктазы не выше, чем в неиммунизированных до вторичного заражения. Активность ФАЛ, однако, увеличивается быстрее в иммунизированных тканях с вторичным заражением С. cucumerinum, а не С. lagenarium. Из всех изученных ферментов только повышение активности пероксидазы четко совпадает с началом иммунизации и увеличением лигнификации иммунизированных растений после вторичного заражения.
У устойчивых огурцов лигнификация быстро возникает после проникания С. cucumerinum и проявляется во многих клетках вокруг места проникания. Лигнификацию наблюдали и у восприимчивых огурцов, зараженных этим патогеном, но реакция задерживалась до момента, когда патоген распространился в ткани, и была слабой и диффузной. Такой же процесс протекает в листьях восприимчивых огурцов, зараженных С. lagenarium. В растениях, иммунизированных заражением С. lagenarium, С. cucumerinum или ВНТ и вторично зараженных С. cucumerinum или С. lagenarium, лигнификация идет более быстро, интенсивно и недиффузно по сравнению с неиммунизированными растениями. В иммунизированных растениях, вторично зараженных С. cucumerinum, она охватывает многочисленные клетки вокруг места заражения, а в растениях, зараженных С. lagenarium, выражена в местах проникания, но особенно — под аппрессориями, где проникание не происходит. Процессы лигнификации более локализованы в иммунизированных растениях, вторично зараженных С. lagenarium, в сравнении с С. cucumerinum. После проявления лигнификации оба вида гриба растут слабо.
Некоторые из недавно появившихся сообщений представляют веские доказательства того, что лигнификация — это механизм устойчивости к заболеванию. Она может ограничивать развитие патогенов с помощью следующих механизмов: а) увеличения механической устойчивости стенок клетки хозяина; б) уменьшения восприимчивости клеточных стенок хозяина к разрушению внеклеточными ферментами; в) ограничения диффузии патотоксинов и питательных веществ и подавления роста патогена в результате действия токсических предшественников лигнина или лигнификации патогена. Сообщения о том, что хитин как компонент клеточных стенок многих грибов может служить индуктором и матриксом для лигнификации, совпадают с данными о предполагаемой роли лигнификации в устойчивости к болезням и иммунизации. Лигнификация может также ограничивать распространение вирусов в растениях. Хаммершмидт первым сообщил о лигнификации мицелия грибов. Мицелий Colletotrichum lagenarium и С. cucumerinum лигнифицировался в присутствии препаратов кониферилового спирта или евгенола, перекиси водорода и пероксидазы из иммунизированных растений. Лигннфикация была подтверждена с помощью окрашивания отмытых в воде и растворителях фрагментов мицелия фтор-глюцинолом-HCl и последующего определения разницы спектров ионизации экстрагированного лигнина. Мицелий обоих грибов не окрашивался фторглюцинолом-НСl при инкубировании с конифериловым спиртом и давал едва различимую реакцию при инкубировании с конифериловым спиртом и перекисью водорода. Свободные радикалы, образовавшиеся в результате действия пероксидазы на конифериловый спирт, обладают высокой реактивностью и сами по себе должны быть токсичными. Образуя ковалентные связи с белковыми и углеводными компонентами клеточной стенки грибов, эти радикалы и возникающие полимеры могут ингибировать рост гриба за счет его лигнификации. Кроме того, гриб может быть связан компонентами стенки хозяина, предупреждающими его развитие в тканях. При лигнификации отмечено заметное снижение чувствительности клеточных стенок хозяина к гидролизу грибными гидролазами.
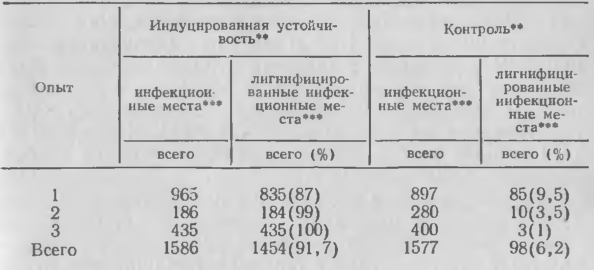
4. Влияние инокулирования грибом С. lagenarium первого настоящего листа промышленного сорта огурца на лигнификацию эпидермиса хозяина в черешках первого листа ниже уровня пазушной почки, вторично зараженной С. cucumerinum
В наших опытах с иммунизированными растениями огурцов образование кониферилового спирта в тканях после вторичного заражения С. cucumerinum или С. lagenarium было нерегулярным. Этот странный факт и заставил предположить, что спирт окисляется пероксидазой сразу же после его синтеза de novo или же образования при высвобождении из растворимых или связанных с клеточной стенкой предшественников. Иммунизированные растения не всегда содержали высокие концентрации растворимых или этерифицированных р-кумариновой или феруловой кислот до или после вторичного заражения. Однако интересно отметить, что конифериловый спирт накапливался в иммунизированных растениях, зараженных С. cucumerinum или С. lagenarium в присутствии восстановителя а-токоферол-ацетата. Конифериловый спирт был высокотоксичным для обоих грибов. В экспериментах на пластинах силикагеля, содержащих 0, 1—0, 3 мкг кониферилового спирта, отмечено явное ингибирование обоих патогенов.
Таким образом, лигнификации может создавать чрезвычайно неблагоприятные условия для грибов, например обусловливая высокую токсичность фенольных предшественников лигнина, токсичность свободных радикалов, лигнификацию грибов, связывание грибов клеточными стенками, повышение устойчивости полисахаридов и белков клеточных стенок хозяина к действию грибных гидролаз. Доказательство роли лигнификации как механизма сдерживания грибов в иммунизированных растениях весомо: грибы попадают в неблагоприятные условия среды, если происходит процесс лигнификации. Исследователи стремятся установить, действительно ли лигнификации ингибирует грибы in vivo и связана ли она с иммунизацией против бактериальных и вирусных болезней. Ограниченное распространение системных заболеваний — вирусных, бактериальных или грибных — можно было бы объяснить действием лигнификации на патогены или на их внеклеточные токсины или ферменты. В иммунизированных растениях после вторичного заражения лигнификации протекает быстрее и с большей интенсивностью. Возникает вопрос — почему это происходит?
Механизмы индукции
До недавнего времени все наши усилия, направленные на иммунизацию растений с помощью абиотических факторов, были безуспешными. Для этого испытывали препараты клеточной стенки грибов и бактерий, их экстракты или внеклеточные продукты, экссудаты флоэмы, экстракты из зараженных и иммунизированных незара-женных тканей. Соответствующими препаратами или экстрактами опрыскивали листья, вводили их в листовые пластинки или вносили в растения через корни, в тест-растворы помещали отдельные листья и всходы. Недавно Гарае и Куч (неопубл. данные) получили активный экстракт из листа-индуктора, зараженного С. lagenarium, ВНТ и P. lachrymans. Введение этого экстракта в семядоли и первый настоящий лист огурца вызвало в расположенных выше листьях иммунитет к заболеванию, вызываемому С. lagenarium. Гесслер и Куч (неопубл. данные) изолировали из листьев огурца, зараженных С. lagenarium, С. cucumerinum, P. lachrymans и ВНТ, белок с молекулярной массой около 16 000 Д, стабильный в кислых буферах и связываемый ДЭАЭ целлюлозой, но не найденный в экстрактах листьев, поврежденных сухим льдом или карборундом. Появление белка совпадает с развитием симптомов болезни, например через 3—4 дня после инокулирования С. lagenarium. Этот белок может быть идентичным тому, о котором сообщили Эндэбран с соавт. Он не найден в тканях системно зараженных растений, но количество белка заметно возрастало в иммунизированных тканях после вюричного заражения, даже если симптомы были слабыми или вообще не появлялись. В иммунизированных листьях после вторичного заражения и в неиммунизированных зараженных листьях с сильными симптомами содержание белка было приблизительно одинаковым. Многие исследователи сообщают об увеличении количества одного или более белков в растениях, зараженных вирусами или обработанных полиакриловой или салициловой кислотой. В настоящее время ведутся эксперименты по выяснению вопроса, является ли белок в зараженных листьях огурца фактором, индуцирующим иммунитет и (или) сдерживающим патоген.
Возможно, что индуктор иммунизации (сигнал) образуется в течение довольно краткого периода после индукции, и этот или другой фактор влияет на нераскрывшиеся листья в точке роста. Нераскрытые листья в соответствующей фазе развития могут быть иммунизированы в течение 4—6 недель. Если через 3 недели провести повторное инокулирование, поражение достаточно молодых, нераскрывшихся листьев обеспечит стабильность иммунитета на весь период плодоношения. Возможность, что какое-то низкомолекулярное транслоцируемое информационное вещество служит компонентом индукции, является в высшей степени спекулятивной, но может быть доказана при наличии соответствующих данных.
Практическое применение
Иммунизация — основа профилактики болезней животных, поэтому удивительно, что она не нашла более широкого применения и в защите растений. Однако данные о практическом использовании иммунизации имеются. В течение многих лет слабовирулентные штаммы ВТМ широко применяют в теплицах и на полях для защиты томатов от вирулентных штаммов вирусов. Круикшанк и Мэндрайк сообщили о защите табака от пероноспороза в поле в результате заражения грибом Peronospora tabacina, хотя это вызывает сильную карликовость растений. Карузо и Куч показали, что слабое заражение огурца, арбуза и дыни грибом С. lagenarium защищает растения в поле от заболеваний, вызываемых при искусственном заражении высокими концентрациями патогена. В некоторых опытах некротизированная зона у иммунизированных растений сокращалась более чем на 98% в сравнении с растениями, которые не были иммунизированы. Контрольные растения, перенесшие заражение, были защищены от антракноза, на рост и урожай иммунизация не влияла (Куч и др., неопубл. данные). Недавно Макинтрай обнаружил, что в полевых условиях табак можно защитить от черной ножки при использовании ВТМ в качестве индуктора.
Использование ВНТ для индукции иммунитета увеличивает технические возможности ее применения в полевых условиях. Раствор, содержащий ВНТ, можно вносить путем опрыскивания под высоким давлением, при этом отпадает необходимость в использовании влажных камер и тщательном контролировании температур для получения ограниченных симптомов. Таким образом, скоро не понадобится индуцировать иммунитет только у всходов в парниках и теплицах и проводить последующую пересадку в грунт.
Иммунизация не обеспечивает защиту растений от всех заболеваний. Нам не удалось иммунизировать тыквенные против мучнистой росы или использовать патоген для индукции. Большинство наших полевых опытов проведено с использованием большой инокуля-ционной нагрузки для вторичного заражения и ограничено определенными климатическими и почвенными условиями. Иммунизация не снижает урожай огурцов, однако табак, иммунизированный против пероноспороза, становится карликовым в отсутствие высокой инсоляции и повышенного уровня питательных веществ. Процент карликовости у иммунизированных растений табака по сравнению с неиммунизированными в условиях слабой интенсивности освещения и при относительно низком содержании питательных веществ в почве варьирует в зависимости от сорта.
Конечно, тот факт, что в растениях развивается какой-то химический защитный барьер, еще не свидетельствует о безопасности растений или растительных продуктов или их пригодности на корм скоту. По мере развития наших исследований мы надеемся выделить и определить химические свойства индуктора иммунизации. Возможно, это вещество будет эффективным при обработке листьев, введении через корни или при протравливании семян.
Основная защита против болезней все еще заключается в получении устойчивых растений селекционным путем. В настоящее время селекция обеспечивает единственный механизм устойчивости к вирусным заболеваниям и экономически обоснованную устойчивость к бактериальным заболеваниям. Мы начали исследования по селекции растений, легко поддающихся иммунизации. Маловероятно, чтобы иммунизация исключила химическую защиту, так же как вакцинация животных не устранила применение антибиотиков — пестицидов животного мира.
Используемая литература: Инфекционные болезни растений: физиологические и биохимические основы/Пер. с англ. Л. Л. Великанова, Л. М. Левкиной, В. П. Прохорова, И. И. Сидоровой; Под ред. и с предисл.
Ю. Т. Дьякова. — М.: ВО Агропромиздат, 1985. — 367 с.
Скачать реферат:
Пароль на архив: privetstudent.com