Развитие симбиотических ассоциаций между бобовыми растениями и клубеньковыми бактериями представляет огромный интерес для агрономии. Эти симбиозы обеспечивают большую часть биологически связанного азота, доступного сельскохозяйственным культурам.
Клубеньковые бактерии — довольно обычные грам-отрицательные микроорганизмы, объединенные в одно семейство с известными фитопатогенными бактериями Agrobacterium. Хотя клубеньковые бактерии рассматриваются в этой статье исключительно с точки зрения их роли в симбиотических взаимоотношениях с растениями, необходимо всегда помнить, что они очень хорошо приспособлены к длительному выживанию в почве даже в отсутствие специфических растений-хозяев. Точно так же и хозяева этих бактерий — бобовые растения, например соя и клевер, могут хорошо расти без своих симбионтов. Таким образом, симбиоз бобовых растений и клубеньковых бактерий — это скорее факультативная, а не облигатная ассоциация. Она обычно выгодна для обоих организмов, но не обязательна для их развития. Каждый партнер этой ассоциации может вести свою собственную жизнь и хорошо для этого приспособлен. Следовательно, можно ожидать, что в процессе инфекции и установления симбиоза развитие клубеньковых бактерий должно быстро изменяться, т. е. происходит адаптация, позволяющая им перейти из комплексной среды обитания в почве в динамическую среду на поверхности корня, а затем в совсем особую среду внутри клеток хозяина. Растение-хозяин должно также быстро реагировать на присутствие симбиотических бактерий на поверхности его корней, причем ответные реакции хозяина должны заметно отличаться от их реакций на другие почвенные микроорганизмы. В растении-хозяине должны произойти значительные внутренние изменения, в результате которых оно сможет принять своего бактериального партнера и переключиться с потребления связанного азота из экзогенных источников на усвоение симбиотически образованного NH3. Совершенно очевидно, что процесс инфекции и развития симбиотических отношений состоит из целой серии мутуалистических адаптаций. Во многих отношениях этот процесс напоминает инфицирование восприимчивого хозяина патогенным организмом, и в то же время они совершенно различны.
Инфицирование сои Rhizobium japonicum, очевидно, начинается с реакции бактерии на появление сигнальных и питательных веществ, выделяемых корнями хозяина. Клетки Rhizobium очень быстро прикрепля-ются к поверхности корня, и их можно обнаружить уже спустя несколько секунд или минут после инокуляции корней. Прикрепление бактерий чаще всего полярное. Следующий этап, наблюдаемый в инфекционном процессе, — скручивание корневых волосков. Скручивание и деформация последних специфически индуцируются клубеньковыми бактериями. Скручивание начинается спустя несколько часов после инокуляции и становится заметным примерно через 12 ч . Клубеньковые бактерии проникают в скрученные корневые волоски у точки наибольшего изгибания и внедряются в них в виде трубчатой структуры, называемой инфекционной нитью. Эти нити переносят клетки ризо-бий, обычно в виде одной цепочки, к основанию корневого волоска (к базальной клетке). Кончик растущей инфекционной нити, по-видимому, движется по пути ядра клетки хозяина. До того момента как инфекционная нить выйдет из базальной клетки волоска, в близлежащих кортикальных клетках происходит стимуляция деления и заполнения цитоплазмой. Инфекционные нити проникают через клеточную стенку, расположенную между клеткой корневого волоска и прилегающими к ней кортикальными клетками, и разветвляются, захватывая многие непрерывно делящиеся кортикальные клетки. Затем происходит высвобождение клеток клубеньковых бактерий из кончиков инфекционных нитей в цитоплазму клеток хозяина. Сразу же после высвобождения бактерии инкапсулируются цитоплазматическими мембранами клеток хозяина и, таким образом, никогда не вступают в прямой контакт с цитоплазмой хозяина. Клетки клубеньковых бактерий и расте-ния-хозяина пролиферируют, формируя структуры, называемые клубеньками, которые состоят из примерно равных в весовом соотношении масс бактерий и тканей растения. Хозяин обеспечивает ризобии продуктами фотосинтеза, а бактерии осуществляют превращение атмосферного N2 в NH3, снабжая растения доступным азотом.
Это удивительный процесс, замечательный по специфике, механизмам действия, развитию и регуляторной утонченности. Наблюдение за ним и изучение его доставляет истинное наслаждение.
Мы начали систематическое и многоплановое изучение инфицирования con Rhizobium japonicum. Моими коллегами в этой работе были физиологи Арвинд Бхаг-ват, Маргарет Пирс и Т. В. Бхуванесвари, цитологи Джиллиан Терджен, Марк Пенс и Гарри Калверт, биохимик Эндрю Морт и специалисты по мутагенезу Ян Ло и Поко Ямамото. Исследования клубеньковых бактерий были начаты с изучения механизмов распознавания и специфичности по отношению к хозяевам. По крайней мере, для большинства бобовых растений, обитающих в зоне умеренного климата, существует четко выраженная специфичность к ризобиям. Например, Rhizobium meli-loti формирует клубеньки только с люцерной и не инфицирует клевер или другие виды бобовых. Штаммы R. japonicum образуют клубеньки лишь с соей, но не с люцерной, клевером и т. д.
Молекулярные основы специфичности хозяев и клубеньковых бактерий
Специфичность хозяина и механизмы распознавания в течение многих лет представляют обширную область исследований для фитопатологов, включая и тех специалистов, которые изучают симбиозы Rhizobium. В 1975 г. Альбершейм и Андерсон-Праути выдвинули теоретическое предположение о том, что в реакции распознавания в системах растение — патоген участвуют связывающие углеводы белки (т. е. лектины) хозяина, реагирующие с углеводами на поверхности клеточных стенок микроорганизмов. Почти одновременно Болул и Шмидт привели экспериментальное доказательство тесной корреляции между возможностью различных клубеньковых бактерий инфицировать сою и их способностью связывать меченый лектин в семенах этого растения. Мы проверили с максимальной тщательностью опыты Болула и Шмидта и подтвердили корреляцию между образованием клубеньков и связыванием лектина ризобиями. Однако в научной литературе отмечены некоторые отклонения от указанной закономерности:
— Бретауэр и Пэкстон, Чен и Филлипс, Ло и Стрийдом привели данные, свидетельствующие об отсутствии корреляции между связыванием лекти-нов и образованием клубеньков у сои;
— некоторые штаммы R. japonicum очень активно образовывали клубеньки, но не связывали лектин сои;
— у ряда штаммов ризобий в культуре лишь небольшое число клеток (1—5%) связывало лектин сои.
На основании наблюдений мы установили одну важную закономерность, а именно способность клеток Rhi-zobium japonicum связывать лектин сои резко меняется с возрастом культуры. Это открытие позволяет предположить, что появление или исчезновение соединений, служащих рецепторами лектина, на поверхности клеток Rhizobium каким-то образом регулируется. Изучив возможность синтеза рецептора лектина в зависимости от веществ, присутствующих в корнях хозяина, мы установили, что все испытанные штаммы R. japonicum связывают лектин сои при выращивании в присутствии корней хозяина, тогда как на лабораторных средах многие из штаммов не имели клеток, способных связывать лектин. Влияние корней на образование рецепторов лектина позволяет объяснить противоречивость в результатах опытов с теми штаммами бактерий, которые образовывали клубеньки, но не связывали лектин, а также понять отсутствие корреляции между этими процессами в опытах других исследователей. Однако нам до сих пор неизвестно, какое из веществ в корнях растения-хозяина ответственно за изменения в синтезе рецептора.
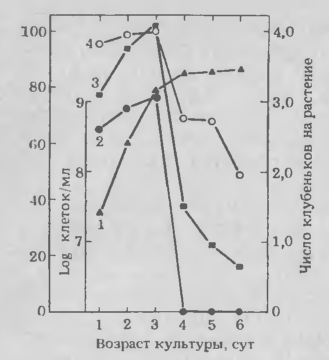
Рис.1. Влияние возраста культуры Rhizobium japo nicum штамм 110 ARS на число клеток, способных связывать лектин сои, и на образование клубеньков на первичных корнях выше отметки исходного положения кончика корня (см. текст) в момент инокуляции:
1 — число клеток/мл; 2 — процент клеток, связывающих лек-тин; 3 — число клубеньков выше отметки в пересчете на растение; 4 — процент растений, имеющих клубеньки выше отметки.
Целесообразно также отметить, что Даззо и Хаббелл установили аналогичную четкую корреляцию между связыванием лектина и образованием клубеньков в симбиотической ассоциации клевер/Rhizobium trifolii.
Стабильность корреляций со связыванием лектина в системе соя/R, japonicum позволяет рассмотреть еще два вопроса:
— можно ли утверждать, что подходящий лектин, находящийся на поверхности корней сои в нужное время и нужном месте, действительно играет роль в реакции узнавания?
—какова природа рецептора лектина сои на поверхности клеток Rhizobium japonicum?
Определенного ответа на первый вопрос в настоящее время пока нет. Лучшим доказательством в его пользу могут быть данные Стейси и др., которые показали, что меченые антитела, специфичные для лектина семян сои, связывались корнями сои в противоположность меченым антителам неспецифической сыворотки. Низкая плотность связывания меченых антител может объяснить причину трудного обнаружения лектина в корнях другими, более прямыми биохимическими методами.
Более полные и точные данные получены по второму вопросу. В соответствии с убедительными цитологическими и биохимическими экспериментами лектин сои связывается материалом капсулы, окружающей клетки Rhizobium japonicum. Меченный ферритином лектин сои связывается капсулярным материалом бактерий, но не внешней мембраной клеток. Эта связь биохимически специфична, что показано методом ингибирования гаптенами сахара, и свободна от влияния артефактов, возникающих при фиксации, как было установлено с помощью криоскалывания.
Мы выделили капсулярный материал у двух различных штаммов R. japonicum и установили, что он состоит из полисахарида, структура которого представлена на рисунке 2. Возможно, что наиболее важной особенностью капсулярного полисахарида (КПС) является наличие боковой галактознльной цепи в каждой повторяющейся единице полимера. Связывание лектина сои поверхностью бактериальной клетки особенно сильно ингибируется при добавлении галактозы или N-ацетнлгалактозамина, показывая, что галактозоподобные структуры составляют важную часть материала, специфически связывающего лектин. Галакто-зильные остатки КПС иногда содержат метильную группу в позиции 4. По данным Хаммарстрема и др., лектин сон не присоединяется к 4-О-метилгалак-тозе. Поэтому особый интерес представляло открытие изменения состава сахаров, входящих в КПС, с возрастом культур бактерий. В старых культурах галакто-зильные остатки в КПС чаще были 4-О-метилированы.
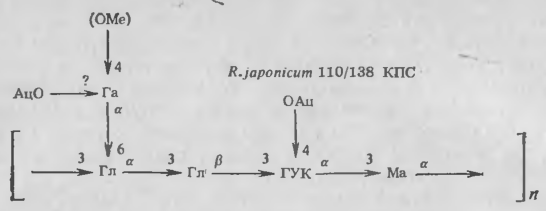
Рис. 2. Структура повторяющейся пентасахаридной единицы, образующей капсулярный полисахарид/экзополисахарид (КПС/ЭПС) в штаммах 110/138 R. japonicum:
Гл — глюкоза; Ма — манноза; ГУК — галактуроновая кислота; Га — галактоза; Ац — ацетат; Me — метнл.
Превращение неметилированной галактозы в метилированную происходит особенно быстро в период, когда бактериальные культуры переходят от экспоненциальной к стационарной фазе роста. В это же время происходит внезапное уменьшение числа инкапсулированных клеток и бактерий, способных связывать лек-тин сон. Изменения в составе КПС и в инкапсуляции объясняют внезапную потерю этими штаммами способности связывать лектин. Как будет показано дальше, при достижении стационарной фазы роста эти штаммы также внезапно теряют инфекционную способность. Пока не установлено, существуют ли какие-либо причинные взаимосвязи между изменениями химического состава КПС, связыванием лектина и инфекционностью Rhizo-bium.
Необходимо учитывать, что штаммы R. japonicum синтезируют как капсулярный полисахарид (КПС), присоединяющийся к хозяину, так и экзополисахарид (ЭПС), не соединяющийся с хозяином. По данным Дад-мена, некоторые штаммы R. japonicum образуют ЭПС, структура которого представлена на рисунке 3. Этот ЭПС, выделенный из культуральных фильтратов, достоверно не содержит галактозоподобных остатков, способных связывать лектин сои. Если последний выступает в роли ви-доспецифичного детерминанта в реакциях распознавания для всех бактерий, образующих клубеньки на сое, следовательно, эти штаммы R. japonicum должны, в свою очередь, также образовывать, по крайней мере в малых количествах, какой-то КПС или ЭПС с сайтом связывания лектина по типу галактозы. По некоторым предварительным данным, два штамма клубеньковых бактерий, которые образуют ЭПС рамнозного типа, способны также синтезировать небольшие количества галактозосодержащих КПС/ЭПС (Морт, неопубл. данные).
Способность клеток R. japonicum связывать лектнн сон строго зависит от наличия капсулярного материала на бактериях. Однако образование клубеньков от этого не зависит. Мутанты R. japonicum, не формировавшие заметных капсул в любой фазе роста как на лабораторных питательных средах, так и при культивировании с корнями хозяина, тем не менее довольно легко образовывали клубеньки на корнях сон. Важным детерминантом способности к образованию клубеньков является возможность синтезировать адекватные количества галактозосодержащего полисахарида. Между количеством образуемого ЭПС, синтезированного различными мутантами ризобий, и их относительной способностью формировать клубеньки существует линейная связь. С появлением некоторого количества галактозосодержащего полисахарида перестает иметь значение, связан ли он с клетками бактерий или находится в свободном состоянии.

Рис. 3. Структура повторяющегося мономера в тетрасахариде, входящем в состав ЭПС штамма 61А76 Rhizobium japonicum; обозначения те же, что и на рисунке 2 (Rha — рафиноза).
1. Взаимосвязь между синтезом ЭПС и формированием клубеньков на корнях сои различными мутантами Rhizobium japonicum, не дающими нормальные капсулы

Можно считать слишком упрощенной идею о том, что лектпны хозяина и полисахариды поверхности клеток Rhizobium — это единственные детерминанты специфичности и распознавания при взаимодействии симбионтов. Если Даззо и Стейси с соавт. считают, что лектнны хозяина ответственны за специфичное прикрепление гомологичных клубеньковых бактерий к поверхности хозяина, то другие исследователи особо выделяют индукцию специфических ответных реакций хозяина с помощью образования связей типа лек-тин — полисахариды. И все же, по-видимому, более вероятно, что эти реакции взаимодействия представляют собой лишь часть сложной последовательности взаимообменов между обоими симбионтами по принципу сигнал— ответ. Согласно этой точке зрения, каждый этап в такой последовательной цепи по-своему определяет общую биологическую специфичность симбиоза. Ни один из этапов процесса распознавания нельзя исключать только потому, что он не является первым или наиболее селективным, и в то же время нельзя считать, что первый или наиболее селективный этап будет полностью определять специфичность симбиотических отношений.
Ранние процессы при инфекциях, вызываемых RHIZOBIUM
Об инициации или раннем развитии инфекций, вызываемых клубеньковыми бактериями у их симбионтов, известно удивительно мало, хотя некоторые последние работы дают по этим вопросам ценную информацию. Как показано ранее, первое внешнее проявление инфекционного процесса — это прикрепление бактерий к поверхности корней хозяина. Клетки Rhizobium, как правило, прикрепляются полярно. По мнению Даз-зо, такое прикрепление определяется лектином хозяина и специфично для системы клевер/R, trifolii. Стейси с соавт. привели аналогичные данные в отношении R. japonicum на корнях сои. Однако мы установили, что и R. trifolii, и R. japonicum могут прикрепляться полярно к корням сои, причем при вполне сравнимых количестве и плотности (Терджен и Бауэр, не-опубл. данные). Характер прикрепления обоих видов практически не различался в условиях наших экспериментов. Независимо от того, является присоединение специфическим или неспецифическим, а иногда промежуточным, приходится признавать, что пока ничего не известно ни о клеточных и молекулярных компонентах, обусловливающих прикрепление, ни о механизмах действия этих компонентов.
Второе проявление инфекционного процесса — индукция скручивания корневых волосков. Функция этого явления заключается, по-видимому, в защемлении клеток Rhizobium в пространстве между двумя поверхностями клеточных стенок изогнутого корневого волоска. До сих пор никому не удалось определить вещества, индуцирующие скручивание или ветвление корневых волосков, не изучены и механизмы, с помощью которых происходит скручивание.
Третье проявление инфекционного процесса — формирование инфекционной нити. Недавние исследования Каллахэма дали очень важную информацию об этом процессе. Он предложил три возможных модели проникания Rhizobium в клетку хозяина:
— локальная инвагинация клеточной стенки корневого волоска;
— локальное разрушение клеточной стенки волоска с помощью индуцированных ферментов самого хозяина и синтез инфекционной нити;
— локальное разрушение клеточной стенки корневого волоска ферментами Rhizobium и синтез инфекционной нити.
Результаты исследований Каллахэма убедительно показали, что процесс инфицирования клевера бактерией R. trifolii включает локальное разрушение клеточных стенок корневых волосков вблизи от захваченных бактерий, связанное с локальным отложением нового слоя неидентифицированного материала, из которого формируется инфекционная нить. Это отложение очень напоминает образование футляра (чехла) вокруг инфицирующих гиф патогенных грибов. Цитологические исследования Каллахэма позволили исключить гипотезу об инвагинации клеточной стенки корневого волоска, однако не дали решения вопроса о том, чьими ферментами вызывается локальное разрушение клеточных стенок — ферментами хозяина или бактерии? Некоторые сообщения поддерживают оба возможных варианта, но не дают определенного ответа.
Множество вопросов, касающихся дальнейших этапов инфекционного процесса, также до сих пор не решено. Например, как может инфекционная нить проникать в клетку хозяина против ее тургорного давления? Почему (и как) кончик растущей инфекционной нити следует за движением ядра в клетке корневого волоска? Как инфекционная нить проникает в соседние кортикальные клетки? Каким образом стимулируется деление кортикальных клеток при развитии инфекционного процесса, вызываемого прониканием нити? Как могут клубеньковые бактерии избегать нормальных защитных реакций растений-хозяев или подавлять их?
К сожалению, серьезные методологические сложности мешают детальному изучению более ранних этапов инфекционного процесса. Сложности возникают из-за того, что в процессе инокуляции обычно участвует около 100 тыс. клеток бактерий и около 10 тыс. корневых волосков, однако инфекционный процесс инициируется не более чем в 100 случаях и лишь около 10 случаев инфекции завершается формированием клубеньков. Поэтому трудно рассмотреть механизмы или неиз-вестные этапы инфекционного процесса в 10 успешных или в целом в 100 случаях инфицирования корневых волосков на фоне 10 тыс. или даже большего количества неудачных или невосприимчивых взаимодействий хозяина и Rhizobium.
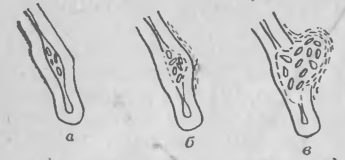
Рис. 4. Диаграмма, показывающая фазы инфекционного процесса, вызываемого клубеньковыми бактериями. Клубеньковые бактерии, заключенные в пространство между двумя поверхностями изогнутого корневого волоска, разрушают клеточную стенку хозяина (а), вызывают синтез нового слоя материала со стороны цитоплазмы клеточной стенки хозяина (б) и проникают через частичио разрушенную клеточную стенку корневого волоска в инфекциониую нить, сформировавшуюся из вновь отложившегося слоя (в).
Локализация инфицируемых клеток на корнях растения-хозяина
Для облегчения методики изучения биохимических механизмов процессов распознавания и инфицирования мы недавно решили определить, какие клетки корня хозяина восприимчивы к инфекции. Проростки растений выращивали в пластиковых пакетах, что позволяло вести визуальное наблюдение за развивающимися корнями. Положение кончиков корней (КК) и зоны образования корневых волосков отмечали на пластике в момент внесения инокулюма. Спустя неделю после инокуляции определяли положение каждого клубенька с учетом сделанных отметок. Если учесть перемещение эпидермальных клеток во время роста корня, можно вычислить положение и фазу развития тех клеток корня, которые были восприимчивы к инфицированию в момент инокуляции клубеньковыми бактериями. Графики (рис. 5. 5) четко показывают, что клубеньки наиболее часто образовывались на тех участках корней, которые находились непосредственно под зоной корневых волосков и над зоной быстрого растяжения. Относительно мало клубеньков формировалось в зоне развивающихся волосков корня, а на участках корней, в момент инокуляции покрытых зрелыми волосками, их вообще не было. Таким образом, способность к формированию клубеньков ограничена определенными фазами развития корней и кратковременностью существования клеток-мишеней, восприимчивых к бактериям. Эксперименты с более поздней инокуляцией сон показали, что весь участок корня выше кончика заметно теряет восприимчивость через 4—6 ч. У других видов бобовых способность к образованию клубеньков также кратковременна и ограничена определенными фазами развития.
Эти данные представляют несомненный теоретический и практический интерес. Например, капельки ино-кулирующей суспензии объемом в нанолитр, содержащие лишь несколько клеток клубеньковых бактерий, можно наносить непосредственно на участок наибольшей восприимчивости корня, и развитие Rhizobium произойдет именно в точке инокуляции. Этот метод позволил нам использовать электронный микроскоп для изучения самых ранних процессов, которые происходят еще до образования инфекционной нити, скручивания корневых волосков или каких-либо иных показателей начала инфекции, заметных в световом микроскопе. Проведение таких исследований возможно только при достаточной уверенности в том, что корневые волоски, взятые для приготовления ультратонких срезов, пригодны для этой цели и будут инфицированы клубеньковыми бактериями.
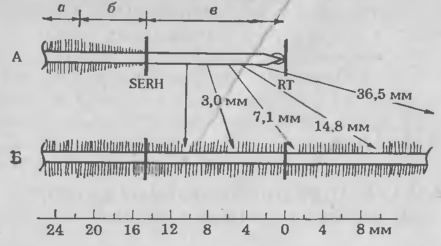
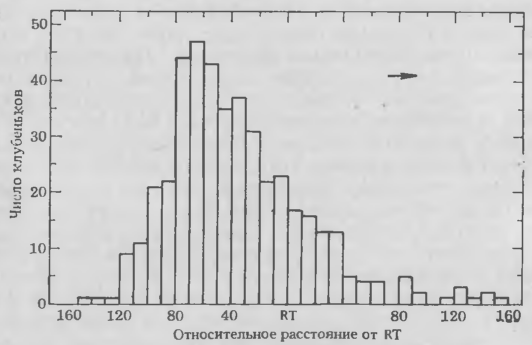
Рис. 5. Частота образования клубеньков в различных зонах корней сои, инокулированных штаммом 110 R. japonicum. Растения выращивали в пластиковых пакетах, на поверхности которых были сделаны отметки, показывающие положение коичика корня (RT) и место наименьшего развития корневых волосков (SERH). Корни приблизительно 120 растений исследовали через неделю после инокуляции, определяя относительное (%) расстояние от каждого клубенька, образовавшегося на первичном корне, до RT. Для более легкого определения относительного положения неинфицированных клеток кория во время инокуляции рассчитано перемещение эпидермальных клеток относительно RT.
Вещества, влияющие на интенсивность начальных процессов инфекции
Не менее интересный аспект исследований темпов развития восприимчивости бобовых растений к клубеньковым бактериям состоит в том, что в настоящее время появилась возможность изучать факторы, влияющие на скорость инициации успешного инфицирования. Число клубеньков, развивающихся над кончиком корня (RT) выше сделанной на пластике отметки, служит прекрасным количественным параметром развития инфекции. Например, Бхагват (личное сообщение) недавно обнаружил, что образование клубеньков некоторыми ризобиями на корнях коровьего гороха происходит непосредственно под меткой, если бактерии культивировали на среде вместе с корнями хозяина перед инокуляцией. Эти результаты позволяют предположить, что Rhizobium могут адаптироваться или отвечать каким-то образом на вещества, выделяемые из корня хозяина, прежде чем они приобретут способность к инфицированию.
У сои можно наблюдать обратное явление. Образование клубеньков на корнях сои выше отметки RT мало зависит от предварительного культивирования R. japonicum в корневых экссудатах хозяина. Однако образование клубеньков с максимальной частотой происходит ниже зоны развития корневых волосков, хотя даже цитологические исследования показали, что инфекция, приводящая к образованию клубеньков, происходит через короткие, только начинающие развиваться корневые волоски. Различия между локализацией ожидаемой и действительной зоны максимального образования клубеньков привели нас к заключению о наличии определенного лаг-периода между временем первого взаимодействия клеток корней с бактериями и временем начала успешного инфекционного процесса в них. Очевидно, этот лаг-период можно сократить, предварительно обработав корни клеточным полисахаридом клубеньковых бактерий. N-ацетилгалактоза, возможный полисахаридный гаптен лектина сон, также эффективен в этом отношении. Такие эксперименты трудны для точного воспроизведения, но нам удалось найти способы, позволяющие контролировать большую часть меняющихся показателей.
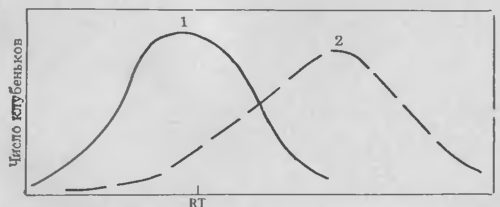
Относительное расстояние между клубеньками и меткой у кончика корня (RT)
Рис.6. Влияние экссудатов коровьего гороха на развитие клубеньковых бактерий:
1 бактерии на экссудатах корней; 2 — бактерии на синтетической питательной среде.
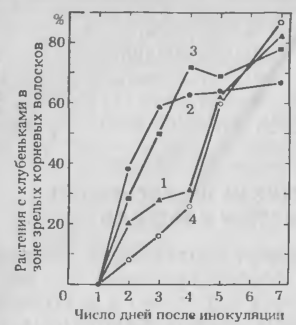
Рис. 7. Влияние бесклеточ-ного фильтрата Rhizobium tri-folii на образование клубеньков в зоне зрелых корневых волосков у клевера. Корни клевера предварительно обработали 100 мкл воды (4) или культурального фильтрата в течение 6, 14 и 24 ч (на графике — 1, 2, 3 соответственно) до внесения инокулюма. Число клубеньков. образовавшихся в зоне корней со зрелыми корневыми волосками в момент инокуляции, определяли ежедневно в течение недели.
Аналогичное явление отмечено и в корнях клевера. Если культуру R. trifolii отмыть, ресуспендировать в дистиллированной воде в течение часа и затем отфильтровать, то свободный от клеток фильтрат заметно сокращает время, необходимое для образования клубеньков в зоне зрелых корневых волосков, при обработке корней этим фильтратом перед инокуляцией. Интересно отметить, что такая же обработка корней клевера не отражалась заметно на времени образования клубеньков в зоне без корневых волосков.
Фильтрат действует на образование клубеньков только в зоне зрелых корневых волосков, очевидно, влияя на второй тип инфекции, характерный для клевера, но не отмеченный у сои, коровьего гороха или люцерны.
Активные соединения, присутствующие в фильтратах суспензий R. trifolii или в корневых экссудатах коровьего гороха, до сих пор не идентифицированы. Тем не менее обнаружение их действия подтверждает точку зрения на то, что взаимное распознавание клубеньковых бактерий и их хозяев представляет комплекс реакций, происходящих между симбионтами по типу сигнал — ответ.
Инфекционная способность клеток RHIZOBIUM
К инфицированию ризобиями восприимчивы только определенные клетки корней бобовых, причем восприимчивость зависит от времени и фазы развития растения. Что же можно сказать о клубеньковых бактериях? Зависит ли инфекционная способность Rhizobium от времени, фазы развития или среды? Влияние корневых экссудатов коровьего гороха на формирование клубеньков ризобиями позволяет предположить, что, по крайней мере, некоторые виды бактерий должны подвергаться определенным изменениям, прежде чем они будут способны вызвать инфекцию. Конечно, даже в одной культуре клетки Rhizobium могут отличаться друг от друга множеством признаков. Например, они могут иметь разные способность к инкапсуляции, подвижность, фазы клеточных циклов, число полярных телец. Возможно, что инфекционная способность зависит от одного или нескольких подобных специфических признаков. Из вышеприведенных данных следует, что в отличие от синтеза ЭПС инкапсуляция сама по себе, видимо, не связана с инфекционной способностью. По данным недавних исследований, инфекционная способность клубеньковых бактерий не зависит от их подвижности.
Работы, проведенные в нашей лаборатории, позволили установить, что инфекционность R. japonicum в значительной мере зависит от возраста культуры. Культуры штаммов 110 и 138 R. japonicum, достигнув фазы стационарного роста, быстро теряли способность вызывать образование клубеньков выше отметки RT. Очевидно, в этой фазе бактерии в меньшей мере готовы или способны вызвать инфекционный процесс во время инокуляции, чем клетки в экспоненциальной фазе. Основа этого изменения к концу экспоненциальной фазы не известна. Изучение влияния дозы инокулюма на процесс образования клубеньков показало, что культуры штамма 110 R. japonicum, находящиеся в ранней или поздней фазе стационарного роста, могут индуцировать образование клубеньков так же быстро и интенсивно, как и культуры в фазе экспоненциального роста, однако для инокуляции растений необходимо большее количество инокулюма старых культур.
Следовательно, различия в инфекционной способности между культурами бактерий в экспоненциальной или стационарной фазе роста являются скорее количественными, чем качественными. Находящиеся в стационарной фазе роста культуры, по-видимому, содержат в пропорциональном отношении меньшее число клеток, способных быстро вызывать инфекцию.
Заслуживает внимания еще один аспект образования симбиотических ассоциаций. Графики частот образования клубеньков показывают ослабление интенсивности этого процесса в нижних участках корней, которые в момент инокуляцин были на 5— 10 ч моложе, чем участки с максимальной активностью образования клубеньков. Очевидно, растения сои обладают быстродействующим регуляторным механизмом, служащим для предупреждения развития излишнего числа клубеньков на корнях. Второе возможное объяснение состоит в том, что убывание числа клубеньков в нижних участках корней может быть связано с уменьшением числа бактерий, приходящихся на долю этих частей. Однако, если эти же корни повторно инокулировали спустя 10—12 ч после первого нанесения бактерий, никаких изменений в образовании клубеньков не происходило, т. е. необходимости в повышении численности бактерий не было. В то же время если первичный инокулюм был нанесен только на верхнюю часть корня без корневых волосков и на зону молодых, развивающихся волосков, где удлинение корня полностью или большей частью завершено, то вторичная инокуляция вызывала формирование новых клубеньков. Этот эксперимент показывает, что быстродействующий регуляторный механизм в растении сон включает в себя некоторые типы взаимодействий между Rhizobium и теми клетками хозяина, которые локализованы в зоне быстрого удлинения корня вблизи от его кончика.
Наличие быстродействующего регуляторного механизма в растениях сои и, возможно, коровьего гороха и люцерны создает дополнительный уровень сложности и организации в динамическом влиянии фаз развития и окружающей среды на раннее развитие и успех инфекций клубеньковыми бактериями.
Используемая литература: Инфекционные болезни растений: физиологические и биохимические основы/Пер. с англ. Л. Л. Великанова, Л. М. Левкиной,
В. П. Прохорова, И. И. Сидоровой; Под ред. и с предисл.
Ю. Т. Дьякова. — М.: ВО Агропромиздат, 1985. — 367 с.
Скачать реферат:
Пароль на архив: privetstudent.com