Симптомы заболевания — это четко выраженное повреждение хозяина, вызываемое патогеном. Обычно симптомы считают визуальным проявлением болезни, но они могут возникать и на других уровнях, например на ультраструктурном или биохимическом. Однако я вынужден признать, что метаболические или даже ультраструктурные симптомы не всегда приводят к визуальному симптому. Для любой болезни обычно характерен не один, а набор визуальных симптомов, появляющихся и параллельно, и последовательно при развитии заболевания. Эта совокупность симптомов, или синдром болезни, как ее обычно называют, достаточно типична, чтобы использоваться для идентификации возбудителя болезни или, по крайней мере, существенно облегчить такое определение, даже несмотря на то, что типы симптомов ограничены количественно и неспецифичны.
Симптомы, появляющиеся на хозяине, можно систематизировать различными способами. Для наших целей точная схема не важна, и вполне достаточно общих закономерностей, которые могут быть выведены из нее. Одна из таких классификаций дана в таблице 1, где я разделил симптомы на два основных типа — визуальные и биохимические, причем каждый из них также подразделяется. По материалам таблицы можно сделать следующие важные выводы: 1) способы, которыми растения могут реагировать на патоген, ограничены количественно; 2) следовательно, ответные реакции растений неспецифичны (например, многие возбудители индуцируют один и тот же симптом); 3) каждый симптом сам по себе является комплексным и в определенной степени может быть связан с другими симптомами.
Схема 1

Факторы, ответственные за возбуждение патогеном симптомов болезни, называют первичными или вторичными детерминантами заболевания, а в последнее время — факторами патогенности и вирулентности. Факторы патогенности являются качественными признаками, а вирулентности — количественными. В особых случаях возникает вопрос о том, с фактором какого типа приходится иметь дело, так как некоторые из них можно отнести одновременно к обоим типам. Например, мутанты Pseudomonas syringae pv. savastanoi, не синтезирующие индолил-3-уксусную кислоту (ИУК) из триптофана, не образуют галлов, хотя по скорости роста в тканях хозяина они аналогичны штамму дикого типа по крайней мере в течение двух недель и формируют галлы при добавлении экзогенного триптофана. Другие мутанты синтезируют повышенные количества ИУК и индуцируют образование более крупных галлов, чем изо-лят дикого типа. В первом случае ИУК можно считать фактором, обусловливающим патогенность (т. е. первичным детерминантом, или фактором патогенности), тогда как во втором случае она связана с вирулентностью (вторичный детерминант, или фактор вирулентности).
Эти факторы, точнее молекулы (токсины, сложные вещества, участвующие в распознавании, ферменты), высокоспецифично взаимодействуют с веществами хозяина. В настоящее время большинство наших исследований сфокусировано на попытке разобраться в этих взаимодействиях, и это вполне понятно. Однако, поскольку мы хотим добиться полного понимания, необходимо двигаться дальше и сконцентрировать все внимание на том, что связывает эти первичные взаимодействия и дальнейшее развитие симптомов заболевания, возникающих в результате этих взаимодействий.
Взаимосвязи между первичными сайтами действия и симптомами болезни
В настоящее время физико-химические взаимодействия, или связи, как я их буду называть, между первичным сайтом действия фактора в хозяине и возникающим симптомом (симптомами) в лучшем случае малопонятны, а обычно вообще не исследованы. Некоторую информацию дает: 1) специфическая реакция (реакции) модельных биологических систем на действие очищенного детерминанта; 2) сравнение с более изученными соединениями, о которых известно, что они действуют в том же сайте; 3) возможность воздействия на развитие симптомов путем вмешательства в биохимические процессы в какой-то точке между первичным сайтом действия и проявлением симптома; 4) познание нормального развития реакций в изучаемой метаболической системе.
Понимание направления процессов, происходящих в период между первичными взаимодействиями и синдромом заболевания, имеет значение не только из-за их научной ценности и потенциальной возможности практического использования. Оно подтверждает первичный механизм действия, предполагаемый для данного детерминанта. Оно дает нам потенциальную возможность не допустить развития симптомов и, возможно, практически контролировать развитие болезни, изменяя направление некоторых реакций, протекающих вне сайтов первичного взаимодействия с паразитом. Оно также позволяет по-новому оценить нормальную жизнедеятельность растений и даже фитопатогенных микроорганизмов.
Исторически сложилось так, что взаимосвязи между детерминантами и симптомами рассматривались в очень упрощенной форме: детерминанты, вызывающие в растении одинаковые симптомы, воздействуют на хозяина в одном и том же сайте и более или менее одинаково. Однако очень скоро, с началом нового этапа в изучении болезней растений, стало ясно, что ситуация является более сложной. Один и тот же симптом болезни может быть вызван патогеном в разных местах. Например, увядание может быть вызвано следующими способами: 1) при нарушении поглощения воды корневой системой; 2) при задержке передвижения воды в сосудистой системе; 3) в результате увеличения потерь воды за счет транспирации. И конечно же, существует множество условий, приводящих к изменениям каждого из этих процессов и в конце концов — к тому же увяданию.
Ситуация еще более усложняется, если учесть, что один или большее число детерминантов может вызывать у растения один и тот же симптом, воздействуя на разные стороны обмена веществ хозяина. Примером может служить симптом хлоротичности. Даже только токсины, индуцирующие хлороз у растений, действуют самыми разнообразными путями: фазеоло-токсин ингибирует фермент орнитин-карбамоилтранс-феразу, влияющий на образование цитрулина, тентоксин подавляет фактор сопряжения хлоропластов (CF1), останавливая, следовательно, фотосинтетическое образование АТФ; ризобитоксин ингибирует β-цистатионазу — фермент, участвующий в биосинтезе метионина, а тагетитоксин нарушает некоторые ранние, независимые от света стадии развития хлоропластов (Люкенс и Дурбин, неопубл. данные). Кроме того, хлороз вызывают и другие токсины, сайты действия которых пока еще не уточнены, но они, вероятно, отличаются от вышеописанных вследствие их биологической специфичности и химического строения. Наконец, помимо токсинов, существуют другие типы детерминантов, вызывающих хлороз, например ферменты. Следовательно, любой патоген образует ряд детерминантов, каждый из которых участвует в возбуждении одного-единственного симптома болезни. Упрощенным примером этого, вновь заимствованным из работ с токсинами, служит синтез одним и тем же изолятом Alternaria alternata нескольких биогенетически не связанных токсинов, вызывающих у растений хлороз с помощью различных механизмов (например, тентоксин, тенуазоновая кислота, альтернуен, альтернариол и его монометиловый эфир). Все это справедливо и для изо-лятов Alternaria mali, токсины которых вызывают некроз.
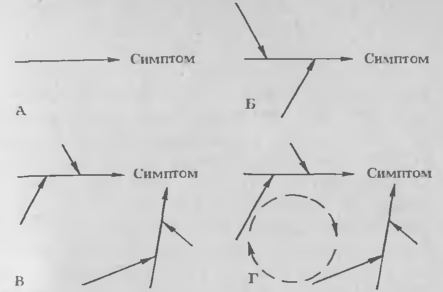
Рис.1. Диаграмма взаимодействия детерминантов патогенности с хозяином, приводящего к появлению симптомов заболевания. Стрелками обозначены физико-химические реакции хозяина, сопровождающиеся симптомами. Детерминанты патогенности действуют на-конце каждой стрелки.
Известны случаи действия детерминантов в сайтах, общие для разных метаболических процессов. В качестве примера можно назвать токсин таб-токсинин-β-лактам. Он подавляет глутамин-синтетазу (ГС), вызывая накопление одного из субстратов этого фермента — аммония (Томас и Дурбин, неопубл. данные). Концентрация аммония достигает в тканях хозяина достаточно токсического уровня, чтобы вызвать разобщение процессов фосфорилирования и нарушение ультраструктуры хлоропластов. Аммоний, накапливающийся в результате ингибирования ГС, образуется при функционировании двух разных путей азотного метаболизма: ассимиляции и фотодыхания. Подавление ГС приводит также к ослаблению синтеза белка и нуклеиновых кислот из-за отсутствия глутамина. Все эти процессы по-разному вызывают нарушение целостности хлоропластов и, следовательно, появление хлороза. Действительно, легко себе представить, что один детерминант, воздействуя на какой-либо сайт, общий для множества метаболических путей, может вызвать два или даже больше симптомов. К тому же один детерминант способен действовать в двух или большем числе определенных сайтов, хотя я предполагаю, что молекулярная среда в этих двух сайтах будет аналогичной, особенно в случае с низкомолекулярными токсинами. Примером может служить ризобитоксин, который, как уже говорилось, ингибирует фермент β-цистатионазу, а также фермент, участвующий в превращении метионина в этилен. В качестве других примеров можно назвать табтоксин, подавляющий ГС и рибулозо-1, 5-дифосфатдекарбоксилазу, и тентоксин ингибирующий CF1 и, возможно, одну или более связанных с этим ферментом реакций. Ультра-структурные наблюдения также позволяют предположить, что токсин AM I Altemaria mali действует в двух сайтах — хлоропластах и плазмалемме хозяина, хотя пока еще не установлено, представляют ли они два биохимически различных сайта или же просто две субклеточные локализации одного и того же сайта.
Некоторые детерминанты не могут вызывать самостоятельно таких реакций, которые привели бы к формированию визуального симптома. Скорее их действие позволяет другому детерминанту влиять таким образом, что симптом возникает как результат этого последовательного действия (как, например, при разрушении клеточной стенки хозяина пектин-метилэстера-зой, которая делает пектин более доступным для разложения его другими ферментами, гидролизующими полисахариды). Еще одним примером может служить взаимодействие молекул, осуществляющих «распознавание» паразитом хозяина; метаболическим следствием их деятельности является возможность протекания других реакций взаимодействия. В то же время некоторые детерминанты в большей степени требуют параллельных реакций, чем последовательных.
Можно себе представить и иной ход событий, включающий одно или большее число превращений детерминанта, прежде чем он сможет вступить во взаимодействие с хозяином и вызвать симптом заболевания. Мы знаем по крайней мере один пример такого типа реакции: табтоксин должен быть гидролизован в растении, чтобы дать табтоксин-β-лактам. Только последняя форма может действовать в первичном сайте хозяина, сам же табтоксин не обладает биологической активностью.
Наибольшее после фитогормонов внимание исследователей было направлено на изучение таких детерминантов, которые разрушают или непосредственно подавляют специфические морфологические структуры или определенные звенья метаболизма хозяина. Другая группа детерминантов, представляющая интерес, включает те из них, которые могут действовать на регуляторные процессы хозяина. Изменения pH, энергетического, водного или окислительно-восстановительного потенциалов на метаболическом уровне оказывают большое влияние и могут приводить к появлению одного или большего числа симптомов болезни через различные обменные процессы. Кроме того, возможны нарушения в процессах транскрипции, трансляции и пост-трансляционных реакциях. Вирусные детерминанты, безусловно, влияют именно на один или несколько таких процессов.
Количественная оценка симптома (симптомов) обычно используется для определения потерь урожая, так как между этими двумя величинами существует положительная корреляция. Однако эта взаимосвязь резко изменяется в случае устойчивых сортов, снижение урожая которых не соответствует активности развития патогена и (или) выражению симптомов. Следовательно, это особый случай, когда знание связи (связей) между детерминантами болезни и симптомами может приобрести огромное значение. Какие же пути метаболизма, позволяющие хозяину стать устойчивым к патогену, могут быть модифицированы и (или) активированы? Данные, полученные при изучении таких защитных реакций, могут найти широкое применение для других систем паразит — хозяин.
Типы детерминантов
Необходимо как можно больше знать о том, как образуются детерминанты болезни и что (в смысле генетических и биохимических механизмов) регулирует их образование. Существуют веские доказательства того, что многие детерминанты являются «адаптивными», т. е. для их образования нужны определенные условия или субстраты. Это сразу же наводит на мысль о возможном использовании регулируемого образования детерминантов в наших интересах. Примером могло бы быть контролирование детоксикации пизатина путем воздействия на образуемый патогеном фермент моно-оксигеназу, ответственную за деметилирование пизатина. Этот тип управления был бы особенно эффективен, если бы его удалось направить против детерминантов, участвующих в первичном нарушении механизма устойчивости хозяина и в процессах распознавания.
До сих пор я говорил о тех детерминантах, которые действуют на хозяина прямым или положительным образом. Но следует рассмотреть и второй тип с противоположным или негативным действием. Это реакции, приводящие к выведению патогеном питательных веществ из тканей хозяина. Определенные качественные и количественные нарушения питания, безусловно, должны вызывать у хозяина нестабильность метаболизма и привести к появлению симптомов заболевания. Лучшим примером этого типа может быть симптом карликовости, вызываемый нематодами или облигатнымн паразитами.
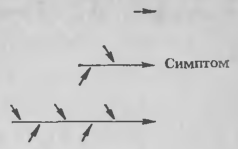
Рис. 2. Схематическое изображение трех степеней (вверху, в середине и внизу) сложности и выраженности физико-химических процессов в тканях растения, влияющих как на детерминанты патогена, так и на симптомы хозяина.
В конечном счете все ответные реакции (т. е. симптомы) хозяина связаны с детерминантами патогенов, хотя связи эти иногда еле заметны. Они могут быть совсем короткими (рис. 2 вверху), как при разрушении клеточной стенки хозяина, промежуточными (рис. 2 в середине) и очень сложными, включающими множество реакций (рис. 2 внизу). В соответствии с этой основной точкой зрения не так уж важно, является ли конечный этап, ведущий к проявлению симптома, результатом действия какого-либо детерминанта патогенности или же следствием образования веществ при первичной реакции самого хозяина, так как в последнем случае детерминанты патогенности все равно участвуют в реакции, но на более ранней стадии. В то же время детерминант не зависит от числа и комплексности реакций, обусловливающих появление симптома, которые он индуцирует. Очевидно, с удлинением связей увеличивается и число детерминантов, необходимых для этого симптома. По существу, все случаи взаимодействий, хотя они могут казаться разными, можно объединить в единую концепцию, при условии что она не противоречит исходным постулатам.
Мне кажется, что логическим, поэтапным анализом связей между детерминантами и симптомами мы сможем постепенно объяснить причины болезни на биохимическом уровне. Выяснение этих связей будет сложной задачей, так как они включают более высокие уровни сложности и взаимодействий, чем наблюдаемые в первичных сайтах действия. И все же вознаграждение за труды будет велико и приведет нас как к пониманию причин возникновения болезни, так и, что еще
более важно, к рациональной разработке мероприятий по защите, которые позволят подавить активность детерминантов болезни.
Используемая литература: Инфекционные болезни растений: физиологические и биохимиче-
ские основы/Пер. с англ. Л. Л. Великанова, Л. М. Левкиной,
В. П. Прохорова, И. И. Сидоровой; Под ред. и с предисл.
Ю. Т. Дьякова. — М.: ВО Агропромиздат, 1985. — 367 с.
Скачать реферат:
Пароль на архив: privetstudent.com