Предварительная инокуляция растений авирулент-ными или несовместимыми расой или штаммом патогена часто дает начало серии биохимических изменений, которые индуцируют в месте взаимодействия устойчивость к вирулентному или совместимому патогену. Широкое изучение молекулярной основы такой индуцированной устойчивости показало, что в проявлении ее участвуют фитоалексины и другие низкомолекулярные соединения, образующиеся в месте взаимодействия. Этот тип динамической реакции наряду с другими физическими и химическими барьерами входит в механизмы защиты, которые растения приобрели в процессе эволюции и которые определяют тот факт, что устойчивость в природе является правилом.
Следовательно, вирулентные или совместимые патогены должны были приобрести способность не только преодолевать статические защитные барьеры, но и подавлять действие генов устойчивости или делать их неэффективными. Однако, за исключением роли специфических токсинов в патогенезе, механизм индуцированной восприимчивости как положительной функции патогенов не исследован столь широко, как индуцированная устойчивость. Давно известно, что растения, предварительно инфицированные вирулентным патогеном, приобретают восприимчивость к слабовирулентным патогенам или непатогенным организмам, и это явление получило название предрасположенности, индуцированной восприимчивости или приобретенной восприимчивости. Подобный тип индуцированной восприимчивости позволяет предполагать, что патогены активно подавляют механизмы защиты хозяина с целью установления псевдосимбиотической ассоциации с клетками хозяина, это и привело нас к концепции доступности.
Ниже приведены некоторые примеры синергизма во взаимодействиях хозяина и паразита в соответствии с современным пониманием индуцированной восприимчивости.
Синергические взаимодействия патогенов в растительных тканях
1. Синергизм при размножении вирусов
При вирусных болезнях взаимодействие патогена и клетки хозяина значительно более прямое, чем при бактериальных и грибных болезнях, и может быть охарактеризовано как геномное. Взаимодействия между разными штаммами или типами вирусов можно подразделить на три группы: 1) нет явного эффекта; 2) антагонистические взаимодействия; 3) синергические взаимодействия. Варьируя условия эксперимента, даже в одной комбинации вирус — хозяин можно наблюдать эти три фазы.
Синергические взаимодействия, которым особенно посвящена эта статья, обнаруживаются на уровне как концентрации вируса, так и проявления симптомов. Полосчатость томатов, вызываемая вирусом табачной мозаики (ВТМ) и вирусом X картофеля (ХВК), морщинистость картофеля, вызываемая ХВК и вирусом А картофеля (АВК), и морщинистая мозаика табака, вызываемая ХВК и вирусом Y картофеля (YBK), — это классические примеры новых симптомов, индуцируемых смешанными инфекциями. Крупные некротические пятна, образующиеся на листьях картофеля при инокуляции ХВК и ВТМ, представляют другой тип синергического эффекта. Локальные пятна на листьях Nicotiana glutinosa или табака NN, образующиеся в ответ на заражение ВТМ, были крупнее при одновременной инокуляции ХВК и содержании в ино-куляте большего количества ВТМ. Если растения томатов инокулировали ХВК и ВТМ, концентрация ХВК была в несколько раз выше, чем в растениях, инокули-рованных только ХВК, в то время как концентрация ВТМ снижалась наполовину или на треть. Концентрация ХВК не возрастала, если инокуляцию ВТМ проводили намного раньше, и это показывает, что метаболические изменения во время размножения ВТМ важны для усиления размножения ХВК.
Концентрация ХВК в листьях табака возрастает также при совместной инокуляции с YBK, т. е. YBK явно способствует системному распространению ХВК при условии, что температура инкубации благоприятна. Системное распространение ХВК усиливается и под влиянием ВТМ, вируса огуречной мозаики (ВОМ, желтый и зеленый штаммы), вируса мозаики белены, американского, но не типового, штамма кольцевой пятнистости табака (ВКПТ), вируса гравировки табака. Считается, что сопровождающий вирус может блокировать механизм ингибирования, который индуцируется в отдельных растениях при инокуляции одним ХВК, и обусловливает перенос ХВК в флоэме.
Дальнейшие опыты по размножению ХВК в листьях табака показали, что синтез патогена усиливается в присутствии YBK или ВТМ в одной и той же клетке . Успешная передача повиликой вируса кольцевой пятнистости настурции и механическая передача P-вируса редьки японской растениям N. glutinosa в присутствии сопровождающего ВОМ — это некоторые примеры определяющей функции вирусов-помощников. По данным Атабекова с соавт., в системе табак — ВТМ температурочувствительный мутант репродуцируется, если к инокулюму добавлен дикий штамм. Эти примеры синергических взаимодействий позволяют предполагать, что вирусы-помощники индуцируют инфекцию или при помощи комплементирующих факторов, необходимых для размножения аберрантного или по иным причинам слабовирулентного вируса, или путем блокировки защитного механизма в отношении этих вирусов.
Наиболее убедительное подтверждение индуцированного совместимого взаимодействия получено при изучении таких инфекций, при которых непатогенные вирусы размножаются в поддающихся определению количествах в листьях, дважды инокулированных вирулентным вирусом. Если листья ячменя инокулировали ВТМ вместе с вирусом штриховатой мозаики ячменя (ВШМЯ) или вирусом мозаики костра (ВМК), размножение ВТМ, в иных случаях очень слабое, заметно усиливалось и становилось системным. По мнению исследователей, это означает возможное влияние вируса-помощника на рост и развитие растения-хозяина, предрасполагающее к системному распространению ВТМ.
Вирусы мозаики люцерны, хлоротической крапчатости вигны, южной мозаики фасоли, некроза табака и желтой мозаики турнепса не распространялись системно в листьях ячменя, одновременно зараженных ВМК. Следовательно, в соответствии с другой интерпретацией этого явления, система репликации, закодированная в геноме вируса-помощника, может поддерживать усиленное размножение ВТМ, но не других вирусов, вероятно, потому что по уровню ограничения они отличаются от вируса-помощника. Вирус-помощник должен по меньшей мере доводить клетки хозяина до состояния, благоприятного для размножения непатогенного вируса, прежде чем обеспечить его системное распространение. Действительно, около 30% протопластов ячменя, ино-кулированных смесью РНК ВТМ и ВМК, флуоресцировало с антителами к ВТМ и ВМК, меченными флуорес-цеин-изотиоцианатом (ФИТЦ), в то время как лишь 1—2% клеток флуоресцировало при обработке антителами к ВТМ при инокуляции только РНК этого вируса. Данный факт — четкое доказательство того, что ВМК действительно функционировал как вирус-помощник при трансляции и репликации РНК ВТМ в листьях ячменя. В будущем роль вирусов-помощников в репликации и проявлении симптомов нужно изучать одновременно с функцией расщепленных геномов многокомпонентных вирусов, чтобы обеспечить подход к программированию вирусных инфекций. При вирусных заболеваниях геномная комплементация вируса и хозяина представляет основу размножения и инфекции. Степени участия генома хозяина в этих процессах следует уделить особое внимание.
Хотя механизм усиленного размножения окончательно не выяснен, можно предполагать, что основой этого типа индуцированной совместимости служат инактивация конститутивных ингибиторов, подавление индуцируемых механизмов устойчивости и комплементиро-ванный или усиленный синтез РНК полимеразы (репли-казы) или ее субъединиц под действием вируса-помощника.
2. Синергическое размножение бактерий в тканях растений
Хорошо известно, что при бактериальных болезнях вирулентность зависит от способности патогенов довести популяцию в ткани хозяина до уровня, достаточного для появления характерных симптомов. Как и при вирусных болезнях, у растений, инокулированных смесью вирулентной и авирулентной или непатогенной бактерий, обычно появляются более слабые признаки поражения, чем при инокуляции одним вирулентным штаммом. В некоторых случаях, однако, заражение смешанным инокулюмом приводит к синергетическому эффекту и сопровождается более выраженными симптомами, чем индивидуальное заражение. При заражении гвоздики Pseudomonas caryophylli и Corynebacterium spp. степень увядания была выше и стеблевая гниль развилась сильнее, чем при раздельной инокуляции. Синергизм наблюдали также при окаймленной пятнистости фасоли. Присутствие Achromobacter sp. в инокулюме привело к образованию большего числа пятен и усиленному размножению Pseudomonas phaseolicola в инокулированных первичных листьях, хотя популяция Achromobacter в листьях не возрастала. Популяционный анализ Xanthomonas oryzae в листьях риса ясно показал, что устойчивые к стрептомицину авирулентные изо-ляты могли размножаться в зараженных листьях, если инокулюм содержал какой-либо вирулентный штамм. О таком же размножении непатогенной Pseudomonas syringae в листьях фасоли, инокулированных смесью P. syringae и P. phaseolicola, сообщил Янг.
Популяции авирулентных штаммов Erwinia amylo-vora в побегах яблони увеличивались при совместной инокуляции с вирулентным штаммом. Авирулентные штаммы Е. amylovora и Е. herbicola защищали восприимчивые побеги яблони от заражения вирулентным штаммом при условии, что антагонисты были добавлены в инокулюм в большом количестве. Однако, когда устойчивый к стрептомицину авирулентный штамм Е. amylovora Е8-5 смешали с вирулентным штаммом в отношении 1: 1 и использовали для инокуляции молодых черешков восприимчивого сорта Джонатан, оба штамма размножались одинаково (рис. 1). Совместимость, определенная по росту Е8-5, была индуцирована штаммами Е9 (вирулентность IV, наивысшая) и 60b (вирулентность I), но не 101b (вирулентность III) того же патогена. Совместимость, индуцированная высоковирулентным штаммом Е9, была эффективна с штаммами Е8-5, 101b и Е. herbicola, но неэффективна с штаммом 60b и другими испытанными бактериями, включая Erwinia stewartii, Pseudomonas solanacearum, P. tabaci, P. syringae, Xanthomonas malvacearum, X. phaseoli и Agrobacterium tumefaciens. Размножение штамма 60b и E. herbicola ингибировалось при смешанной инфекции с вирулентным штаммом Е9. Эти результаты показывают, что индукция совместимости в данной системе достаточно сложна. Так как интерференция между штаммами Е. amylovora в культуре отсутствовала, снабжение питанием за счет вирулентных бактерий, как часто предполагалось, едва ли служило основным механизмом индуцированной совместимости (Оучи с соавт., неопубл. данные).
3. Доступность при грибных болезнях
После первого сообщения о повышении восприимчивости к желтой ржавчине у пшеницы, зараженной твердой головней, индуцированная восприимчивость была описана для многих комбинаций растений и грибов, особенно при болезнях, вызываемых облигатными паразитами. Такая биологическая предрасположенность была абсолютной в одних комбинациях, но относительной в других, и лишь в немногих случаях обсуждались ее возможные механизмы. Так как сапрофиты способны расти в мертвых тканях, заболевания, вызываемые некротрофами, были отнесены к непригодным для анализа индуцированной восприимчивости. Действительно, наиболее убедительны работы, проведенные с болезнями, вызываемыми биотрофами. Ниже приведены некоторые примеры исследований, тесно связанных или направленных на раскрытие механизма индуцированной восприимчивости.

Рис. 1. Размножение вирулентных и авирулентных штаммов Е. amylovora в тканях черешков, инокулированных простым или смешанным инокулюмом. Для инокуляции черешков молодых побегов яблони Джонатан использовали суспензию клеток 108 клеток/мл стрептомицинустойчн-вого авирулентиого штамма Е8-5 и вирулентного Е9. Число клеток Е8-5 определяли путем посева го-могената зараженных черешков на глюкозодрожжевой агар с добавлением стрептомицина и хлорида трифенилтетразола (ТТХ) с последующим подсчетом колоний: 1, 2 — Е9 при простой и смешанной инокуляции; 3, 4 — Е8-5 при простой и смешанной инокуляцни соответственно.
Хиура при гибридизации специализированных форм Erysiphe graminis первым отметил, что у некоторых гибридов белый мицелиальный мат развивался без образования клейстотециев, что указывало на возможную роль индуцированной восприимчивости во взаимодействии при мучнистой росе. Моузмен и Грили и Моузмен с соавт. показали затем, что возбудители мучнистой росы ячменя и пшеницы образуют конидии на своих нехозяевах, если листья предварительно инокулированы совместимой расой.
Тсучия и Хирата провели опыты с другими мучнистыми росами и обнаружили, что 45 из 51 мучнисторосяного гриба (49 — с двудольных растений) способны инфицировать уже зараженные мучнистой росой листья ячменя, а 30 грибов образуют конидии на таких предрасположенных листьях. Подобный тип индуциро-ванной восприимчивости приняли за основной физиологический процесс, необходимый для патогенеза, и возникла концепция доступности. Доступность, индуцированная заражением совместимой расой Е. graminis f. sp. hordei, была эффективна не только для несовместимых рас этого гриба или других мучнисторосяных грибов, например Е. graminis f. sp. tritici и Spae-rotheca fuliginea, но и для некротрофных непатогенов, например для Mycosphaerella melonis, Glomerella cin-gulata, Colletotrichum lagenarium и Gloeosporium oliva-rum.
Некоторые другие некротрофы не могли образовывать гифы в клетках с индуцированной доступностью, это привело нас к предположению, что общее клеточное истощение вряд ли может быть основным механизмом доступности. Такое предположение было подкреплено данными Тсучия и Хирата, показавшими, что Erysiphe polygoni, собранная на растениях Schizopepon bryoniae-folius Mazim. и Pilea mongolica Wedd, образует конидии, E. polygoni на Fagopyrum esculentum Moench. растет, не образуя конидий, в то время как Е. polygoni с растений Polygonum aviculare L. не может инфицировать листья ячменя, зараженные совместимой расой Е. graminis f. sp. hordei.
В то же время листья ячменя, инокулированные несовместимой расой, оказались недоступными для исходно совместимой расы возбудителя мучнистой росы ячменя. Для завершения клеточных процессов, ведущих к доступности, потребовалось 15—18 ч, а для процессов, ведущих к недоступности, — лишь 6 ч. Обе индуцированные реакции локализованы вблизи места первичного взаимодействия, по крайней мере в ранней фазе инфекции [45, 49], но распространялись и на другую сторону листьев в более поздней стадии. Недоступность в адаксиальном эпидермисе листьев ячменя проявлялась через 18 ч после инокуляции абаксиальной поверхности несовместимой расой и сохранялась лишь 48 ч. Аналогично доступность в адаксиальном эпидермисе проявилась после 72-часового совместимого взаимодействия и достигла максимума на 5—7-й день после первичной инокуляции. Это состояние доступности сохранялось не менее 8 дней после достижения максимума. Таким образом следует отметить, что индуцированная доступность на адаксиальной поверхности была ограничена зоной, немного большей, чем доступная зона на абаксиальной поверхности, инокулированной совместимой расой.
Микроскопическое наблюдение за реакцией живых эпидермальных клеток ячменя на расы Е. graminis подсказывает путь, каким устанавливается доступность для инфекции. Известно, что протоплазматические агрегаты образуются под кончиком аппрессория к моменту внедрения, через 10—11 ч после инокуляции, и исчезают с появлением гаустории, обычно через 2—3 ч после начала ее развития. Временной ход процесса заражения ячменя мучнистой росой обобщен в таблице 1. Это упрощенная схема, представляющая процессы, которые происходят в течение 24 ч совместимого и несовместимого взаимодействий. Следует заметить, что некоторые пропагулы совместимой расы не формировали гаустории, а некоторые несовместимые успешно образовали функционирующую гаусторию. Трудно точно определить отдельные процессы, которые ведут к такому успеху или неудаче, поскольку реакции весьма непостоянны и даже поведение клеток под недоразвитыми аппрессори-ями варьирует от отсутствия заметного противодействия до образования папиллы нормальных размеров. Тем не менее микроскопическое исследование внутреннего эпидермиса колеоптилей ячменя, инокулированных конидиями Е. graminis f. sp. hordei и E. graminis f. sp. tritici, показывает, что успешное или неудачное становление совместимых и несовместимых комбинации может определяться скоростью, степенью, продолжительностью и, вероятно, временем инициации цитоплазматических агрегатов. Роль папилл в защитных механизмах остается предметом споров, и все же папиллы могут развиваться достаточно рано, чтобы функционировать как барьер при быстром и экстенсивном образовании цитоплазматических агрегатов. Агрегаты исчезают или рассеиваются, когда гриб образует функционирующую гаусторию, но сохраняются до тех пор, пока не образуются крупные папиллы, если гриб не способен к образованию гаустории.
1. Временное течение первичных взаимодействий при мучнистой росе ячменя

Следовательно, совместимый или даже образующий гаусторию несовместимый гриб могут иметь некие механизмы, замедляющие или подавляющие образование агрегатов, вероятно, путем нарушения присущей клеткам способности распознавать чужеродные тела. Поэтому совместимые расы должны: 1) избавиться от действия конститутивных соединений: 2) внедриться до того, как образуется папилла высокой плотности, укрепленная флуоресцирующими соединениями; 3) подавить агрегацию цитоплазмы; 4) подавить синтез фитоалексинов, быть нечувствительными к ним или разлагать фитоалексины и 5) подавить гибель мезофилла.
Процессы в клетке, ведущие к ее доступному или недоступному состоянию, необратимы. Эксперименты с тройной инокуляцией показали, что ставшие доступными клетки не распознаются даже непатогенами, например грибом с дыни, как несовместимые. Наоборот, листья, у которых была индуцирована недоступность, при последующем взаимодействии с совместимой расой реагировали более обширным некрозом, чем при взаимодействии с несовместимой расой.
Подобная же необратимость распознавания отмечена при корончатой ржавчине овса. Проявление устойчивости листьев овса к несовместимой расе Puccinia соrо-nata f. sp. avenae ослабевало при обработке совместимой расой в пределах 8 ч несовместимого взаимодействия. Однако заметного эффекта не было, если несовместимое взаимодействие продолжалось 12 ч или более перед инокуляцией совместимой расой. Листья овса с индуцированной устойчивостью, т. е. после 12 ч несовместимого взаимодействия, распознавали инокулн-рованную затем совместимую расу как несовместимую, что привело к усиленной реакции устойчивости. Эти результаты четко показывают, что совместимая раса обладает способностью, как у мучнисторосяных, подавлять этот индуцируемый механизм устойчивости, прежде чем он достигнет необратимого состояния. Предварительная инокуляция листьев овса совместимой расой возбудителя корончатой ржавчины действительно индуцировала доступность для несовместимой расы. Эта индуцированная доступность эффективна по отношению к непатогенам овса, таким как Uromyces alopecuri и Puccinia gramitiis f. sp. tritici, но не к более близкому P. coronata f. sp. festucae (Тани, личное сообщение). Проявление устойчивости требует активации информационной РНК (иРНК), синтеза белка и фитоалексина, поэтому совместимая раса для установления псевдосимбиотической ассоциации с клетками хозяина должна подавлять действие генных систем путем нарушения распознавания или последующего процесса стабилизации с помощью пока неизвестного механизма.
Индукция доступности не ограничена только болезнями, вызываемыми облигатными паразитами. Листья риса, инокулированные совместимой расой Pyricularia oryzae, становились доступными для несовместимой расы после 18 ч совместимого взаимодействия. Аналогично листовые влагалища риса, инокулированные совместимой расой возбудителя пирикуляриоза, становились доступными для несовместимой расы и непатоге-на Alternaria kikuchiana.
Молекулярные основы индуцированной доступности
Из описанных выше наблюдений можно сделать вывод, что совместимые патогены образуют молекулы, подавляющие или нарушающие присущую клеткам хозяина способность распознавать и соответственно отторгать последовательно внедряющиеся несовместимые патогены. Общее свойство всех патогенов — сдерживать защитные механизмы растений-хозяев. По способам подавления защитных механизмов патогены можно подразделить на три группы: 1) пассивно преодолевающие защитные реакции путем нарушения процесса распознавания за счет таких продуктов, как капсулы или внеклеточные полисахариды; 2) активно подавляющие механизмы устойчивости путем образования токсинов или ферментов; 3) активно подавляющие де-репрессивное действие генов устойчивости путем образования соединений, которые не вызывают заметных повреждений клеток хозяина. Далее приведены примеры, доказывающие значимость этих видов патогенных продуктов в патогенезе.
1. Токсины
Токсины, образуемые патогенами, играют значительную роль в возбуждении и (или) развитии болезней растений и животных. Эти продукты могут быть и специфичными, и неспецифичными, но все они тем или иным путем используются для подавления защитных реакций хозяина. Так называемые специфичные токсины в большинстве случаев представляют собой первичные детерминанты болезни (патотокспны), и их присутствие в конидиальных суспензиях сопровождается колонизацией авирулентными штаммами, изолятами или непатогенами листьев, генетически восприимчивых к продуценту токсина. Например, авирулент-ный штамм Helmitithosporium victoriae (непатоген), Н. саrbonum и даже сапрофит Neurospora crassa заселяли листья сортов овса, чувствительных к HV-токсину, если его добавляли к суспензии конидий этих грибов, хотя кривые роста сапрофита несколько отличались от других. Аналогично авирулентные изоляты A. kiku-chiana приобретали способность инфицировать чувствительные к АК-токсину листья груши при его добавлении к споровой суспензии авирулентных изолятов. Опыты показали, что листья груши, предварительно инокулированные непатогеном и затем — вирулентным изолятом, образуют меньше пятен, чем листья без предварительной инокуляции. Следовательно, листья груши могут иметь потенциал устойчивости к непатоге-нам, и АК-токсин служит химическим средством подавления способности клеток хозяина устранять непатоге-ны. Механизм этого действия не выяснен, но образование токсина при прорастании конидий на ранней стадии инфекции может частично нарушать функции мембраны и таким образом играть роль в процессе становления инфекции. Исходя из этих данных Нисимура с соавт. предположили, что патогенность Alternaria alternata представлена двумя формами: потенциалом агрессивности (способностью к внедрению), общим для всех патотипов вида, и специфической вирулентностью, проявляющейся в образовании одного из специфических для хозяина токсинов. В противоположность некоторым специфическим токсинам, действующим как явные детерминанты патогенности, токсин расы Т Helmi-nthosporium maydis, вероятно, играет несколько иную роль. Генетический анализ рас О и Т этого гриба однозначно показал, что Т-токсин не нужен для установления совместимых отношений, но способствует вирулентности расы Т И. maydis.
Таким образом, роль токсинов представляется многоплановой; одни действуют как фактор патогенности, другие — как фактор вирулентности, что повышает вредоносность болезни.
2. Фактор, ингибирующий сверхчувствительность, и супрессоры
Сверхчувствительная гибель клеток представляет одно из наиболее характерных проявлений устойчивости растительных тканей к несовместимым патогенам и широко изучена при некоторых бактериальных и грибных заболеваниях. Типичный пример — система картофель— Phytophthora infestans. Предварительная инокуляция тканей картофеля совместимой расой приводит к подавлению реакции сверхчувствительности. Это явление недавно было объяснено как процесс, включающий в себя взаимодействия между молекулами хозяина и гриба. Компоненты стенки гиф и зооспор совместимой или несовместимой расы не только связывались мембранной фракцией гомогенатов клубней картофеля, но также индуцировали типичную сверхчувствительную гибель клеток и накопление фитоалексина. Показано, что расосортовая специфичность состоит в подавлении связывания этих компонентов, сверхчувствительной гибели клеток, а также накопления фитоалексинов фактором, ингибирующим сверхчувствительность (супрессором) и выделенным из зооспор, мицелия или жидкости, в которой прорастали споры совместимой расы. Супрессор — это глюкан из 17—23 глюкозных единиц с β-1-3- и β-1-6-связями.
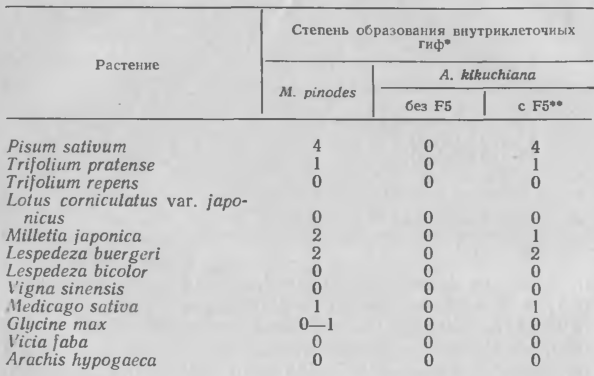
Сходный тип фактора патогенности обнаружен у возбудителя аскохитоза гороха. Хорошо известно, что растения гороха образуют пизатин в ответ на заражение и другие типы стресса. В жидкости прорастающих спор Му-cosphaerella pinodes, возбудителя аскохитоза, обнаружены высокомолекулярный элиситор и низкомолекулярный супрессор образования пизатина. Элиситор, по-видимому, является глюканом с приблизительной молекулярной массой 7Х104Д. Два нингидринположитель-ных вещества, обнаруженных при тонкослойной хроматографии на целлюлозе, подавляют активность элиситора, блокируя накопление пизатина в тканях. Эти два супрессора были предварительно названы F2 и F5. Из двух соединений F5 более активен как супрессор накопления фитоалексина и индуктор доступности для Stemphylium sarcinaeforme, непатогенного для растений гороха (табл. 2), и поэтому использован в качестве модельного супрессора в описанных ниже исследованиях. При содержании в конидиальной суспензии 5 мкг/мл F5 листья гороха становились доступными для непатогенов 5. sarcinaeforme (возб. пятнистости клевера), A. kikuchiana (возб. черной пятнистости груши), Mycosphaerella ligulicola (возб. пятнистости хризантемы), Ascochyta punctata (возб. аскохитозной листовой пятнистости вики) и Cochliobolus miyabeanus (возб. гельминтоспориоза листьев риса). Два первых гриба образовали конидии на обработанных F5 листьях гороха через 14 дней после инокуляции.
Затем специфичность F5 испытывали при инокуляции различных бобовых суспензий A. kikuchiana 15В, содержавшей 50 мкг/мл F5; штамм 15В не образует токсина. Результаты приведены в таблице 3. В присутствии F5 A. kikuchiana приобретала способность заражать те растения, на которых паразитирует образующий F5 гриб М. pinodes, т. е. Pisutn sativum, Trifolium pratense, Milletia japonica, Lespedeza buergeri и Medi-cago sativa.
Таким образом, биологическая активность F5 настолько специфична, что специфичность М. pinodes для хозяина можно объяснить образованием этого продукта конидий. F5, вероятно, подавляет общую защитную реакцию и функционирует как детерминант патогенности. Это соединение не вызывает каких-либо видимых повреждений листьев, стеблей гороха или протопластов из листьев гороха, поэтому его вряд ли можно рассматривать как токсин. Проназа снижала способность F5 подавлять накопление пизатина в ответ на заражение 5. sarcinaeforme, т. е. F5, видимо, содержит пептид. Анализ амннокислот в кислотном гидролизате F5, очищенном жидкостной хроматографией под высоким давлением, показал присутствие аспарагиновой кислоты серина и валина в отношении 1: 2: 1. Таким образом, олигопептид составляет, по-видимому, основную часть молекулы F5. Сходный тип супрессора обнаружен также в жидкости проросших конидий Mycosphaerella melonis. Нингидринположительное соединение из жидкости вызывало доступность семядолей огурца, но не листьев гороха или хризантемы, к A. kikuchiana 15В. Таким образом, биологическая активность этого вещества также соответствует кругу растений-хозяев М. melonis, его продуцента.
2. Влияние супрессоров F2 и F5 на накопление пизатина и развитие гриба в листьях гороха, иноку лированных S. sarcinaeforme

3. Поражение различных бобовых растений М. pinodes и A. kikuchiana 15В при обработке и без обработки растений соединением F5, образуемым М. pinodes
Супрессоры с аналогичной активностью обнаружены также в жидкости прорастающих конидий возбудителя пирикуляриоза риса P. oryzae и уредоспор возбудителя корончатой ржавчины овса P. coronata f. sp. avenae. Внутренние эпидермальные клетки листовых влагалищ риса реагировали типичной сверхчувствительной гибелью на несовместимую расу P. oryzae, но обработка низкомолекулярной фракцией жидкости конидий гриба вызывала внутриклеточный рост несовместимой расы без сверхчувствительной агглютинации цитоплазмы. Листовые влагалища, обработанные той же низкомолекулярной фракцией, становились доступными для A. kikuchiana. Подобным же образом фракция, полученная из уредоспор P. coronata f. sp. avenae, индуцировала доступность листьев овса с различным генетическим фоном. Несовместимая раса росла так, как если бы она была совместимой. Однако при этом листья овса не были доступны для непатогенных ржавчинных грибов, например P. graminis f. sp. tritici, P. coronata f. sp. festucae и P. coronata lolii (Тани, личное сообщение).
Недавние исследования в нашей лаборатории показали наличие низкомолекулярного индуктора в листьях ячменя, зараженных совместимой расой Е. graminis f. sp. hordei. Частично термолабильный низкомолекулярный компонент был обнаружен также в зараженных ржавчиной листьях фасоли. Он способствовал образованию гаусторий Uromyces phaseoli var. typica в листьях вигны и U. phaseoli var. vignae в листьях фасоли.
Все эти наблюдения свидетельствуют о том, что патогены независимо от того, являются они биотрофами или некротрофами, образуют определенные препятствия, которые активно или пассивно противодействуют динамическим защитным реакциям тканей хозяина, для того чтобы обеспечить начало и успешное развитие патогенеза.
Используемая литература: Инфекционные болезни растений: физиологические и биохимические основы/Пер. с англ. Л. Л. Великанова, Л. М. Левкиной, В. П. Прохорова, И. И. Сидоровой; Под ред. и с предисл.
Ю. Т. Дьякова. — М.: ВО Агропромиздат, 1985. — 367 с.
Скачать реферат:
Пароль на архив: privetstudent.com