При поражениях ржавчиной и мучнистой росой термин «сверхчувствительность» применяют в тех случаях, когда некроз клеток хозяина (или нехозяина) связан с устойчивостью независимо от его протекания во времени или от последствий некроза. Поскольку такой некроз при указанных болезнях обычно сопровождает устойчивость, сверхчувствительность следует рассматривать в пределах целого ряда реакций устойчивости. Ранние цитологические исследования тканей, пораженных ржавчиной и мучнистой росой, показали, что некроз клетки хозяина связан с ограничением роста гриба. По-видимому, индуцированная сверхчувствительная гибель клеток ограничивает рост гриба или в результате меньшего доступа биотрофного паразита к живым клеткам хозяина, или за счет выделения веществ, подавляющих или убивающих гриб. Однако в последние годы результаты количественных цитологических исследований, показавших, что подавление гриба и развитие некроза в тканях хозяина не всегда тесно коррелируют, поставили под вопрос значимость гибели клеток хозяина для ограничения роста патогена. Более того, при изучении различных комбинаций хозяин — паразит обнаружено беспорядочное разнообразие во взаимосвязях между некрозами хозяев и подавлением грибов. Цель данной статьи — обзор основных схем сверхчувствительности, обнаруженной в ответ на заражение ржавчинными и мучнисторосяными грибами, и обсуждение предположений относительно причин и последствий сверхчувствительной гибели клеток. В первую очередь сверхчувствительность рассматривается у нехозяев, т. е. видов, иммунных к заражению видами или специализированными формами грибов, и, во вторую — у хозяев, т. е. видов, которые могут быть поражены видами или специализированными формами грибов, но у которых существует расоспецифическая несовместимость, обусловленная взаимодействием по типу «ген на ген».
Сверхчувствительность у нехозяев
Стэкмен ввел термин «сверхчувствительность» в статье, описывающей иммунитет листьев злаков-нехо-зяев к несоответствующим специализированным формам Puccinia graminis. Эти грибы, по-видимому, проникают в ткани нехозяина обычным путем, но затем убивают клетки, расположенные в непосредственной близости. Этот эффект схож (но более ярко выражен) с индуцированным некрозом клеток хозяина при проявлении расоспецифической несовместимости с соответствующей специализированной формой, который позднее также был назван Стэкменом сверхчувствительным. Дальнейшие исследования реакций нехозяев в общем подтвердили наблюдения Стэкмена, показав, что сверхчувствительная гибель клеток нехозяина обычно происходит перед или одновременно с образованием первичной гаустории в месте инфекции, так что вторичные гаустории образуются редко.
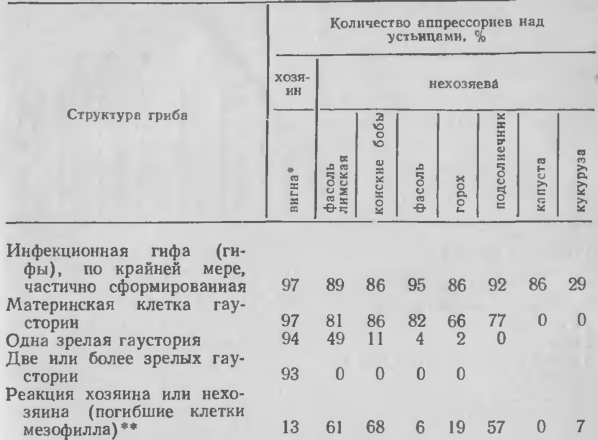
Известно также, что ржавчинные и мучнисторосяные грибы на нехозяевах часто с трудом завершают формирование инфекционных структур перед образованием гаусторий и прекращают рост до индукции реакции сверхчувствительности. Например, по данным Хит (табл. 1), возбудитель ржавчины вигны на различных нехозяевах в большинстве случаев образует мало гаусторий или не образует их вообще, на некоторых нехозяевах не формируются материнские клетки гаусторий и процент мест заражения с погибшими клетками у нехозяев обычно низок. Сходные результаты Хит получила с возбудителями ржавчины подсолнечника и кукурузы. Некоторые нехозяева эффективно препятствовали развитию гриба из материнских клеток гаусторий, и Хит тесно связывает накопление кремнийсодержащих отложений в точке внедрения с неспецифическим механизмом устойчивости у фасоли. В тканях капусты действуют другие механизмы, которые ингибируют скорость формирования инфекционных структур по сравнению с условиями in vitro. Характер развития ржавчинного гриба на разных нехозяевах, по данным Рагги и Тани с соавт., подобным же образом отражает различную степень сопротивления на ранних стадиях инфекционного процесса. Несомненно, что нехо-зяин имеет несколько способов защиты от ржавчинных грибов, которые могут действовать до реакции сверхчувствительности.
1. Развитие возбудителя ржавчины вигны Uromyces phaseoli var. vignae на листьях нехозяев
Мучнисторосяные грибы также редко образуют гаустории в клетках нехозяина. Джонсон с соавт. обнаружили, что Erysiphe graminis f. sp. hordei плохо прорастает на представителях семейств Iridaceae и Li-liaceae и замедленно образует инфекционные выросты на различных нехозяевах из Gramineae (табл. 2). Лишь небольшая часть популяции гриба развивается в достаточной степени, для того чтобы индуцировать сверхчувствительную гибель клеток (табл. 2). Аналогичные результаты получены и с Erysiphe cichoracea-rum. Однако для ростковых трубок, образующих инфекционный вырост, вырост и гаусторию или вырост, гаусторию и гифы, вероятность гибели клеток высока (0, 7— 1, 0) (табл. 2). Следовательно, реакция сверхчувствительности может представлять собой эффективную «последнюю линию» обороны.
2. Развитие возбудителя мучнистой росы E. graminis f. sp. hordei на листьях хозяина и нехозяев
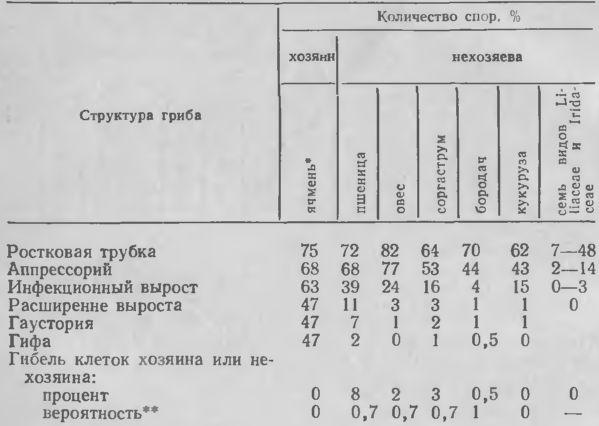
Сверхчувствительная гибель клеток, очевидно, может быть индуцирована у нехозяина и без образования первичной гаустории. Хит предупреждала, что некоторые гаустории в погибших клетках могут остаться незамеченными, и все же ее данные показывают, что клетки обычно отмирают до образования гаусторий ржавчинного гриба, а в некоторых случаях (как с кукурузой) — до образования материнских клеток гаусторий. В опытах с Е. graminis f. sp. hordei клетки нехозяина погибали только после образования инфекционного выроста, хотя так и не установлено, проникает ли вырост через их стенки. Для решения этого вопроса необходимо больше данных, но ржавчинные грибы, вероятно, могут индуцировать сверхчувствительную гибель клеток нехозяина, не проникая в них.
При образовании гаустории клетки нехозяина редко избегают сверхчувствительной гибели, если только не происходит ее инкапсуляции; в последнем случае внедряющуюся гаусторию покрывают вещества, которые, очевидно, отделяют ее от протопласта нехозяина. Если инкапсуляции не происходит, клетка хозяина обычно обречена на гибель. Хит удалось с помощью теплового шока (а в некоторых вариантах при обработке бластицидином S или циклогексимидом) увеличить процент ростковых трубок, способных образовывать гаустории в клетках нехозяев, но эти клетки погибали, если инкапсуляция гаустории не происходила. При некоторых обработках комбинаций нехозяин — гриб патоген продолжал медленно расти в течение многих дней, что сопровождалось побурением и быстрой гибелью пораженных клеток аналогично недетерминированной сверхчувствительности, речь о которой пойдет дальше.
Таким образом, сверхчувствительная реакция нехо-зяина обычно наблюдается в тех случаях, когда нападающий ржавчинный или мучнисторосяный гриб преодолевает защитные барьеры, особенно если образуется неинкапсулированная гаустория. Можно предполагать, что вещества клеточной стенки гриба индуцируют реакцию, как это постулируется для других болезней, однако в настоящее время причина сверхчувствительности нехозяев остается неизвестной. Сверхчувствительность нехозяев заслуживает дополнительных исследований по двум причинам: 1) это в достаточной мере решающая реакция, которая наступает на ранних стадиях инфекции и обычно останавливает дальнейшее развитие гриба; 2) сверхчувствительность — проявление общей, неспецифической способности высших растений противостоять нападению микроорганизмов. Совместимый патоген должен утратить запускающий реакцию механизм или каким-то образом нарушить последовательность биохимических процессов, приводящих к гибели клетки.
Сверхчувствительность у хозяев
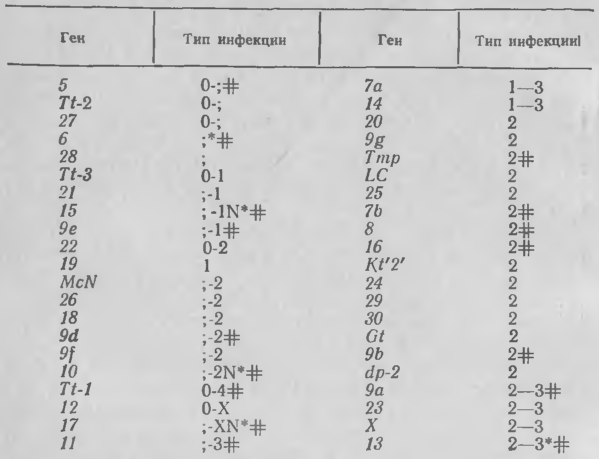
В противоположность нехозяину, в тканях которого быстрое подавление роста гриба почти или совсем не оставляет заметных невооруженным глазом следов, столкновение между соответствующим видом хозяина и несовместимой расой ржавчинного или мучнисторосяного гриба приводит к образованию выраженных участков хлоротичной или некротической ткани. Количество вовлеченной в реакцию ткани и степень роста гриба могут варьировать в зависимости от гена, определяющего несовместимость. Так, в растениях пшеницы с генами Sr устойчивости к стеблевой ржавчине (таблица 3) реакции варьируют от нулевого типа (отсутствие внешних признаков) до мелких пятен (тип; ) и далее до пятен или колец хлоротичных и некротизированных клеток, которые могут достигать нескольких миллиметров в диаметре (тип инфекции 2). Различимый рост гриба происходит при инфекциях типа 1—2. Хлороз обычно более заметен, чем некроз, количественную оценку которого нельзя дать визуально. Однако при типе 1, как правило, развивается заметный некроз, и гены Sr10, Sr15 и Sr17 могут обусловливать образование четко выраженных некротических пятен.
Каждый ген Sr дает несколько типов инфекции, характерных для него (табл. 3). Этот ряд может быть узким, как у Sr5, Sr9е или Sr9b, или широким, как у Sr10, Sr11 или Sr14. Очевидно, каждый ген обусловливает несовместимость своим особым, уникальным путем. Однако весь набор типов инфекции представляет собой коитиниум с большими сходствами, чем различиями. Наиболее распространена неопределенная (недетерминированная) сверхчувствительность, при которой некроз клеток хозяина сочетается с ограниченным, но продолжающимся ростом гриба. Рассмотрим сначала этот недетерминированный тип и позднее перейдем к детерминированному, при котором рост гриба прекращается.
3. Диапазон типов инфекции, обусловленных 42 генами Sr устойчивости к стеблевой ржавчине пшеницы
Недетерминированная сверхчквствительность
Недетерминированный характер роста ржавчинного гриба и сопровождающего его некроза хозяина иллюстрирует таблица 4, по Брауну с соавт. Рост гриба замедлялся к 48 часам, но продолжался в течение 48— 120 ч после инокуляции. Степень ингибирования роста гриба не полностью коррелировала со степенью некро-тизации клеток хозяина как через 48 ч, так и через 120 ч. В целом чем слабее был рост гриба, тем выше был процент некротизированных тканей, но с существенными исключениями. Например, в растениях линии W2402 с Sr9b рост гриба ослабевал при отсутствии некрозов у хозяина. Браун с соавт. пришли к выводу, что сверхчувствительный некроз ткани хозяина не является причиной устойчивости.
4. Зоны заселенной патогеном и некротической ткани у сортов и линий пшеницы с различными генами устойчивости после инокуляции расой 21 Anz-2 P. graminis f. sp. tritici при 18, 5 °С

По данным многих авторов, ржавчинные грибы часто продолжают медленно расти после не-кротизации тканей несовместимого хозяина. Это же отмечено для мучнисторосяных грибов, которые также могут медленно расти после гибели клеток хозяина. Следовательно, гриб остается живым, что подтверждено также методом трансплантации, при которой он рос из несовместимых тканей в совместимые.
Длинные гифы, обычно формирующиеся по краям колонии возбудителя стеблевой ржавчины пшеницы на листьях совместимых сортов, отсутствуют на листьях несовместимых хозяев (Дж. Б. Роуэлл, личное сообщение). В несовместимых тканях эти гифы укорочены, извилисты, а их кончики могут быть вздутыми. Такие особенности роста могут оказаться потенциально полезными как индикатор при поиске веществ, задерживающих рост гриба при недетерминированных реакциях сверхчувствительности.
Чтобы выяснить взаимосвязь между развитием некроза хозяина и ингибированием гриба, Самборскн и его коллеги в Лаборатории сельского хозяйства Канады (Виннипег) детально изучили цитологию недетерминированной несовместимости со стеблевой ржавчиной пшеницы, обусловленной геном Sr6. Они разработали методику с использованием каль-кофтора в качестве фторохрома для определения положения гиф ржавчинного гриба в препаратах целых листьев с помощью светового микроскопа. При низких температурах, когда несовместимость, определяемая Sr6, выражена, они установили, что гаустории образуются через 16—20 ч после инокуляции, некоторые клетки некротизируются через 24 ч и через 72 ч в 85% зон поражения развивается одна или больше некротических клеток. Рост колоний шел значительно медленнее в несовместимом хозяине (Sr6), чем в почти изогенном совместимом хозяине (sr6). Роринджер с соавт. выявили различия в росте гриба на линиях с Sr6 и sr6, через 24 ч после инокуляции. Следовательно, торможение роста начиналось почти одновременно с некротизацией клеток хозяина. При Sr6 рост варьировал: одни колонии прекращали расти, другие медленно росли еще в течение 84—156 ч после инокуляции. С помощью электронной микроскопии Хардер с соавт. обнаружили мертвые гаустории в здоровых и живые гаустории — в погибших клетках хозяина, хотя обычно мертвые гаустории и клетки хозяина находили вместе. Авторы пришли к выводу, что ни некротизированные клетки хозяина, ни некротизированные гаустории «по-видимому, не являются причиной некроза своего партнера по взаимодействию». Результаты показывают, что некроз клеток хозяина, вероятно, может содействовать ограничению роста гриба, но не исключает его подавления другими факторами.
Растения с геном Sr6 теплочувствительны, т. е. теряют устойчивость при 25 °С и выше и дают тип инфекции 3—4. Этот сдвиг в реакции использовали Дейли с соавт. для выявления корреляции между биохимическими процессами и сверхчувствительными реакциями. Например, они показали, что активность пероксидазы оставалась высокой и при 20°С, когда сверхчувствительность была выражена, и при 26 °С, когда сверхчувствительность не проявилась. Казалось бы, это элиминирует роль пероксидазы в сверхчувствительной устойчивости. Однако некоторые признаки несовместимости сохранялись и при 26 °С. Например, число зон поражения составляло 50—60% от числа, полученного при 20 °С; число флуоресцирующих сайтов (использованных как индикатор некротизированных зон) было достаточно высоким, скорость роста гриба при 26 °С на растениях с Sr6 была гораздо слабее, чем на растениях с sr6. Очевидно, листья с Sr6 остаются частично устойчивыми и не могут быть использованы для полной характеристики поведения восприимчивых тканей.
Развитие некрозов и задержка роста гриба, описанные для растений с Sr6 (при 20 °С) и стеблевой ржавчины пшеницы, достаточно хорошо отражают недетерминированную сверхчувствительность. Однако корреляция между обоими процессами не всегда является такой тесной, как показывают данные таблицы 4, и в зависимости от присутствующего гена рост гриба не всегда ослабевает так быстро после инокуляции. Роринджер с соавт. показали, что при наличии Sr8 или Sr22 ограничение роста начиналось через 3—4 дня после инокуляции, когда развился некроз клеток. Для недетерминированной сверхчувствительности характерны продолжающийся медленный рост гриба и прогрессирующий некроз клеток хозяина — неопределенная последовательность гетерогенных процессов, которую крайне трудно исследовать и объяснить.
Детерминированная сверхчувствительная реакция ведет к полной остановке роста гриба. При каждом заболевании ржавчиной или мучнистой росой, для которого установлена модель роста гриба, детерминированная реакция может быть обусловлена одним или большим числом генов (табл. 5). Как подчеркивалось ранее, такие гены значительно менее распространены, чем те, которые контролируют недетерминированную сверхчувствительность. Например, для стеблевой ржавчины пшеницы Sr5 — это единственный изученный ген, который обычно дает детерминированную реакцию, причем лишь на некоторых генетических фонах (Дж. Б. Роуэлл, личное сообщение). Если детерминированная реакция проявляется рано, она очень похожа на сверхчувствительную реакцию нехозяина; первая клетка с внедрившейся гаусторией погибает, и гриб прекращает рост. Гриб может быть убит, это показали Литтлфилд и Ааронсон на примере гена L устойчивости к ржавчине льна, когда патоген терял способность расти из трансплантата в совместимые ткани.
Тани с соавт. изучали детерминированную реакцию в опытах с корончатой ржавчиной овса (Sho-kan 1 — раса 226), при которой рост гриба останавливался через 35 ч после инокуляции. К этому времени около 90% зон поражения содержало одну или более погибших клеток хозяина. Этому предшествовал период (20—35 ч после инокуляции), в течение которого рост гриба ослабевал на 1/3 по сравнению с совместимой комбинацией хозяин — паразит. Следовательно, гриб был мягко подавлен до сверхчувствительной гибели клеток и полностью — после ее проявления. По данным Тани с соавт., при 4-часовой обработке бласти-цидином S, пуромицином или кордицепином через 8— 12 ч после инокуляции несовместимость в системе Sho-kan 1 — раса 226 ослабевала. Это показывает, что триггерные процессы, ведущие к несовместимости, происходят через 8—12 ч после инокуляции или раньше и что синтез РНК и белка необходим для начала процессов, которые позднее заканчиваются внезапным некрозом и остановкой роста гриба.
5. Примеры, детерминированной сверхчувствительности при заболеваниях ржавчиной и мучнистой росой

Детерминированная реакция сверхчувствительности, обусловленная Ml-а геном устойчивости ячменя к мучнистой росе, исследована нами как в интактных листьях, так и в эпидермальных тканях, частично изолированных из колеоптилей. В тканях хозяина с Ml-а большая часть активной авирулентной популяции гриба прекращает рост на стадии папиллы, до образования гаустории, и не индуцирует гибель клеток хозяина. По данным Джонсона с соавт., ген Мl-а усиливает реакцию папиллы. Однако ростковые гифы, не остановленные в папилле (те, которые прекращают рост в отсутствие папиллы или же образуют гаусторию или гаусторию и гифу), с вероятностью 0, 49 индуцируют гибель клеток при поражении в течение 48 ч после инокуляции и с вероятностью -1, 0 индуцируют гибель клеток через 96 ч (Л. Э. Б. Джонсон, неопубл. данные). Гриб не рос после гибели клетки хозяина. Немногочисленные клетки погибали до образования гаустории, но после формирования инфекционного выроста. Таким образом, ген Ml-а, по-видимому, обусловливает две самостоятельные реакции - образование папиллы и сверхчувствительную гибель клеток. Ростковые гифы, прошедшие барьер папиллы, фактически не могут избежать сверхчувствительной реакции.
Нам удалось наблюдать реакцию сверхчувствительности в живых эпидермальных тканях из колеоптилей. В этом случае высокий процент ростковых гиф индуцирует гибель клеток, поскольку лишь немногие из них. задерживаются на стадии папиллы. В целом 40—55% спор, помещенных на ткани, образовало гаустории и 50—100% клеток, содержавших гаустории, погибало через 17—36 ч после инокуляции, когда первичные гаустории сформировались, по крайней мере, частично. В соответствии с более ранними наблюдениями мы нашли, что движение цитоплазмы внезапно прекращалось примерно за 2 ч до гибели клеток хозяина. Ход процессов показан на рисунке 1. Анализ 13 последовательных кадров, полученных при центрайферной съемке, показал, что остановке движения цитоплазмы предшествует период ее усиленного, но локализованного вблизи гаустории перемещения. В течение этого периода отсутствует нормальное перемещение органелл в тяжах. Это отклоняющееся от нормы движение начинается в среднем за 23 мин до остановки и продолжается 17 мин.
Рис. 1. Типичная последовательность процессов при реакции сверхчувствительности эпидермальных клеток ячменя на Е. grami-nis f. sp. hordei; несовместимость обусловлена геном Ml-а.

Неподвижная цитоплазма располагается на клеточных стенках в виде глобул и телец неправильной формы и размеров, но не кажется дегенерирующей. Гибель клетки хозяина происходит внезапно, за несколько минут ей предшествует разрыв ядра (в тех случаях, когда оно хорошо видно). Обычно клетка хозяина гибнет незадолго до гибели гаустории, но в некоторых клетках сначала гибнут гаустории. По-видимому, мембраны гаустории и ядро, плазмалемма и, вероятно, тонопласт хозяина повреждаются примерно в одно и то же время.
Гибель не была ограничена только клетками, содержащими гаустории, часто погибала одна или несколько соседних клеток. В соответствии с исследованиями соседние клетки погибают раньше клеток, содержащих гаустории, и движение цитоплазмы также сначала прекращается в соседних клетках.
Приведенные наблюдения детерминированной сверхчувствительности, обусловленной Ml-а геном, позволяют предположить, что некий токсин, действующий на клетку с гаусторией и на соседние с ней клетки, останавливает движение цитоплазмы и позднее вызывает общее повреждение мембран в клетках хозяина и в гаустории. Интересно, что токсин викторин вызывает очень схожую последовательность процессов: неподвижность цитоплазмы, за которой следует гибель клетки (У. Р. Бушнелл, неопубл. данные). Однако пока еще не установлено состояние клеточных мембран в период остановки цитоплазмы. Мы знаем лишь, что клетка, по крайней мере отчасти, сохраняет тургор до своей гибели.
Время распознавания
Процессы распознавания, которые обусловливают несовместимость, должны происходить до проявления ее первых признаков. Так, детерминированная несовместимость, определяемая геном Ml-а ячменя по отношению к мучнистой росе, выражается в остановке движения цитоплазмы уже через 5—6 ч после начала формирования гаусторий. Как и у нехозяев, некроз клеток хозяев происходит или без образования гаустории, или вскоре после него (см. выше). Однако распознавание должно начинаться задолго до образования гаустории, это показано на примере детерминированной системы Shokan 1 — раса 226 корончатой ржавчины овса, в которой период 8—16 ч после инокуляции был принят за время распознавания (см. выше). Даже при недетерминированной сверхчувствительности распознавание может происходить значительно раньше, как в опытах с Sr6 и стеблевой ржавчиной пшеницы, где некроз проявляется вскоре после образования первичной гаустории. В тех случаях, когда проявление несовместимости задерживается и происходит через 3 или 4 дня после инокуляции, например в системе Sr8 или Sr22 — стеблевая ржавчина пшеницы, раннее распознавание также возможно. Во всех случаях время распознавания должно быть точно установлено. Методы Тани с соавт. с использованием ингибиторов синтеза РНК и. белка представляются перспективными для этой цели.
РОЛЬ гаустории в сверхчувствительности
Первичная гаустория, по-видимому, играет ключевую-роль в реакциях сверхчувствительности, если принять во внимание, что процессы распознавания, вероятно, могут предшествовать ее образованию. Гаустории обычно формируются еще до проявления сверхчувствительности (за исключением некоторых опытов с нехозяевами и с ростковыми гифами у мучнистой росы ячменя, которые не развиваются далее инфекционного выроста). Если сверхчувствительную гибель клеток у линий с теплочувствительными генами задержать с помощью изменений температуры, некротизирующиеся клетки остаются у края колонии, вблизи вновь образующихся гаусторий. Следовательно, зона между стенкой гаустории и протопластом хозяина может представлять участок, где протекают процессы распознавания. Природа веществ, участвующих в распознавании, неизвестна.
Повреждение или нарушение функций в зоне взаимодействия гаустория — хозяин может играть роль в последующей сверхчувствительной гибели клеток хозяина. Хардер с соавт. обнаружили редкие разрывы в экстрагаусториальной мембране (происходящей из ин-вагинированной плазмалеммы) при обусловленной геном Sr6 несовместимости в системе пшеница — стеблевая ржавчина. При таком состоянии, вероятно, может начаться цепь процессов, ведущих к гибели клетки хозяина, или нарушиться перенос питательных веществ от клетки хозяина к гаустории. В тканях, сверхчувствительно реагирующих на заражение ржавчиной, внутри экстрагаусториального матрикса заметны необычные электронно-плотные отложения, которые, возможно, мешают транспорту питательных веществ. В несовместимых хозяевах меченые аминокислоты и другие вещества не способны перемещаться в гифы или гаустории ржавчинных и мучнисторосяных грибов. Такие результаты показывают, что гаустория функционирует не полностью, но не позволяют установить, является ли нарушение функции причиной или следствием сверхчувствительности.
Мучнисторосяные грибы вызывают изменения клеточных мембран у совместимых хозяев. Эти изменения малоизучены, но можно предположить, что они способствуют установлению зависимых биотрофных взаимоотношений между паразитом и клетками хозяина. Например, в экстрагаусториальной мембране нет АТФа-зы нормальной плазмалеммы хозяина. Исследова-ния, начатые нами недавно вместе с О. и Э. Стадель-маннами, показали, что плазмалемма тканей ячменя, зараженных совместимой расой Е. graminis f. sp. hor-dei, плотно прилегает к клеточной стенке, что вызывает вогнутую форму плазмолиза вместо нормальной выпуклой. Это изменение проявилось на следующий день после инокуляции, по крайней мере при высокой плотности колоний. Изменение формы плазмолиза не было следствием изменения осмотического потенциала, который при заражении лишь немного снизился. Вероятно, заражение вызвало изменения в мембране, которые повлияли на степень ее прикрепления к клеточной стенке хозяина. Неспособность индуцировать это или другие изменения, благоприятные для гриба, может играть определенную роль в развитии реакции сверхчувствительности.
Причина гибели клеток хозяина
Ни для одной сверхчувствительной реакции, вызываемой ржавчинными или мучнисторос яными грибами, не удалось установить причину гибели клеток хозяина (или нехозяина). За рядом исключений, например в работе Пруски с соавт., гриб обычно погибает позже, чем клетки хозяина, оставаясь живым неопределенно долгое время при недетерминированной сверхчувствительности или погибая примерно одновременно с клетками хозяина при детерминированной сверхчувствительности. Следовательно, гибель клеток хозянна в целом нельзя приписать действию продуктов погибшего или погибающего гриба. Наоборот, взаимодействие живых хозяина и паразита приводит к образованию токсических веществ, которые могут разрушать клетки хозяина. Токсические вещества могут действовать на расстоянии в несколько клеток от клеток, содержащих гаустории, что обычно характерно для недетерминированной сверхчувствительности. Действие на расстоянии не зависит от гибели пораженной клетки, так как соседняя клетка может погибнуть первой (как в случае с геном Ml-а устойчивости ячменя к мучнистой росе) или не погибнуть вообще. Последний случай наблюдается при наличии гена Sг5 в системе стеблевая ржавчина — пшеница, если гаустория внедряется в эпидермальную клетку. Пораженная клетка остается живой, в то время как соседняя клетка мезофилла погибает (Дж. Б. Роуэлл, личное сообщение).
Самое прямое доказательство роли тожсина в сверхчувствительности к ржавчине или мучнистой росе привел Олиен, использовавший электрический градиент по длине инфицированных листьев пшеницы и получивший смещение сверхчувствительных некротических пятен по направлению к положительному полюсу. Опыт проводился с сортом пшеницы Khapli и расой 56 P. graminis f. sp. tritici (вероятный эффективный ген Sr7a), которая дает тип инфекции 1 с хорошо заметными некрозами через 8—10 дней после инокуляции. Когда рост гриба прекращался, мелкие некротические пятныш-ки сливались в одно пятно диаметром около 1, 5 мм. Если электрический градиент применяли на пятый день, незадолго перед максимальным проявлением некроза, пятно перемещалось на 1—2 мм, так что пустула и пятно теряли концентричность. Таким путем Олиен установил роль отрицательно заряженного токсического вещества как причины некроза. Несомненно, для установления роли токсина в сверхчувствительной гибели клеток необходим его прямой поиск.
Подавление роста гриба
Помимо гибели клеток хозяина, подавлению роста гриба при сверхчувствительных реакциях на заражение ржавчиной и мучнистой росой потенциально способствуют и другие факторы. Фитоалексины участвуют в этом процессе при обеих болезнях. Кроме того, гриб, вероятно, неспособен индуцировать необходимые для оптимального роста изменения в тканях хозяина, как это происходит у совместимых хозяев. Мы уже подчеркивали, что не только сверхчувствительная гибель клеток, но и иные механизмы часто ограничивают развитие инфекционных структур в тканях нехозяина; часть этих механизмов может продолжать действовать независимо от гибели клеток хозяина.
Тем не менее сверхчувствительная гибель клеток увеличивает препятствия для паразита и иногда может быть основным фактором, ограничивающим его рост, особенно если детерминированная сверхчувствительная реакция происходит вскоре после образования первичной гаустории. Микрохирургические эксперименты показали, что при потере первичной гаустории рост Е. graminis продолжается лишь в следовых количествах и гаустория не выживает в погибшей клетке хозяина, если в ней не поддерживать искусственно осмотические условия. Итак, можно допустить, что гриб неспособен расти, если погибает первичная клетка хозяина. Ржавчинные грибы обладают способностью ограниченно ветвиться в тканях хозяина, не образуя гаусторий. И все же гибель первично инфицированной клетки не только лишает гриб функционального взаимодействия с протопластом хозяина, но может привести к выделению из погибшей, декомпартментализованной клетки хозяина ингибирующих веществ.
При недетерминированной сверхчувствительности отрицательные последствия гибели клеток хозяина не столь решающи, но они, вероятно, способствуют задержке роста гриба. Недетерминированные системы заметно реагируют на экспериментальные воздействия. Например, развитие гриба в таких системах можно изменить с помощью разнообразных обработок, особенно таких, которые повышают или снижают содержание растворимых сахаров или аминокислот в тканях хозяина. Иногда рост гриба можно продолжить при наличии некроза у хозяина или остановить в его отсутствие. Такие результаты следует интерпретировать с учетом того, что гриб обычно продолжает развиваться при недетерминированной сверхчувствительности. Следовательно, его рост может быть модифицирован без устранения несовместимости между хозяином и паразитом.
Используемая литература: Инфекционные болезни растений: физиологические и биохимические основы/Пер. с англ. Л. Л. Великанова, Л. М. Левкиной, В. П. Прохорова, И. И. Сидоровой; Под ред. и с предисл.
Ю. Т. Дьякова. — М.: ВО Агропромиздат, 1985. — 367 с.
Скачать реферат:
Пароль на архив: privetstudent.com