Обработка сточных вод и сжигание мусора на судах
Полисомы
Уже давно известно, что рибосомы в цитоплазме клетки часто образуют скопления разной величины, иногда приобретающие удлиненную конфигурацию. Когда появилась гипотеза посредника, было высказано предположение, что рибосомы в группах соединены нитями матричной РНК и, следовательно, являются тем основным участком, где происходит синтез белка в клетке. Мысль о том, что рибосомы в этих группах соединены друг с другом нитью мРНК, основывается главным образом на следующем наблюдении: когда препарат обрабатывали РНКазой в низких концентрациях, эти группы распадались. На некоторых электронных микрофотографиях изолированных полисом также как будто видно, что рибосомы соединены чем-то вроде нити. Поскольку биохимическими методами такую нить в цитоплазме клетки выявить не удалось, возникает вопрос, действительно ли рибосомы соединены РНК.
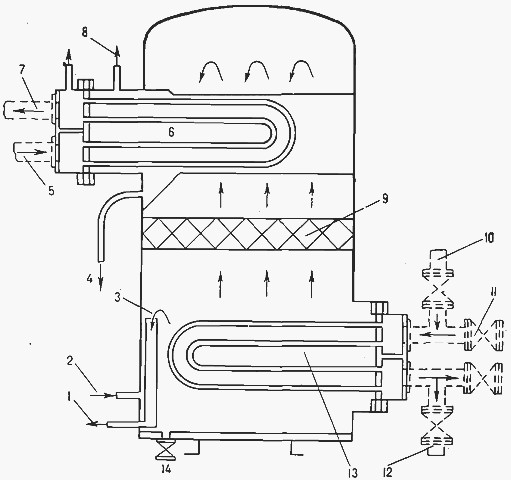
Опреснительные установки
В процессе опреснения морская вода сначала превращается в пар, а затем конденсируется, в результате чего получается пресная вода. Парообразование может происходить при кипении воды при обычном давлении либо при пониженном давлении, когда кипение воды происходит при температуре, меньшей 100 °С. При испарении происходит снижение количества растворенных в воде веществ с 32 000 мг/л до 1—2 мг/л. Аппарат для опреснения называется опреснителем, а иногда дистиллятором.
Процесс кипения. Нагрев воды в испарителях этого типа осуществляется от подогревающих витков трубопровода при пониженном давлении в корпусе испарителя по сравнению с атмосферным. При этих условиях температура кипения воды будет 60°С. Вода к испарителю забирается в месте отлива циркуляционной забортной воды за борт и сначала пропускается через конденсатор, а затем часть ее отбирается для испарительной камеры (рис. 7.6)
Категория: Рефераты / Производство
РНК в цитоплазме клетки
Если некоторые быстрометящиеся ядерные РНК служат «посредниками» переносящими информацию от генов в цитоплазму, то их, очевидно, следовало бы искать не только в ядре, но и в цитоплазме. Настойчивые поиски, которые в течение многих лет с помощью разных методов предпринимали во многих лабораториях, оказались безуспешными: в цитоплазме животных клеток не удалось обнаружить никаких фракций РНК, подобных быстрометящейся ядерной РНК . Первое указание на то, что в цитоплазме может совсем не содержаться таких фракций РНК, было получено при исследовании седиментационных свойств препаратов всей цитоплазматической РНК, экстрагированной фенолом и детергентом. При центрифугировании в градиенте плотности сахарозы в препаратах не оказалось фракций, подобных фракции быстрометящейся ядериой РНК с такой же полидисперсностью или аналогичными кинетическими свойствами. Когда стало ясно, что различное поведение быстрометящейся РНК при седиментации связано с ее чувствительностью к изменениям ионной силы раствора и к содержанию в нем катионов, цитоплазматическую РНК начали исследовать в растворах с разной ионной силой и разными концентрациями катионов; тем не менее в цитоплазме так и не обнаружили РНК с физическими свойствами быстрометящейся ядерной РНК. Наконец, решили использовать колонки с метилированным альбумином, хорошо отделяющие быстрометящуюся РНК от гетерогенной смеси других рибонуклеиновых кислот. Но опять не было обнаружено никакой РНК со свойствами, характерными для быстрометящейся ядерной РНК.
РНК в ядре клетки
Когда клетки животных в течение 10—15 мин инкубируют с радиоактивным предшественником РНК, более чем 90% метки обнаруживается в ядре. Тот факт, что радиоактивные предшественники прежде всего включаются в РНК, отмечен еще в 1948 году Бергстрандом, и с тех пор связь между первоначальным появлением метки в ядре и последующим проникновением ее в цитоплазму стала объектом обширной и сложной литературы. Подобное явление наблюдается и у бактерий. Когда бактерии, имеющие хорошо выраженную ядерную зону, например Е. coli, инкубируют в течение 1 мин с радиоактивным предшественником РНК, большая часть метки оказывается в «ядре», а при более длительной экспозиции метка появляется и в цитоплазме бактериальной клетки. В животных клетках быстрометящаяся ядерная РНК находится не в «растворе», а, по-видимому, присоединена к хромосомам. Способ присоединения новообразованной РНК пока еще не известен: возможно, что около ДНК-матрицы ее удерживают водородные связи, однако есть основания предполагать, что белок также принимает в этом участие. Присоединена ли быстрометящаяся РНК к хромосомам бактерий, пока еще не известно. Для того чтобы выделить эту РНК, клетки обычно разрушают и полученный препарат обрабатывают ДНКазой с целью уменьшения его вязкости. Такая обработка, конечно, разрушает бактериальную хромосому и поэтому лишает смысла любое рассуждение о том, было или не было что-либо присоединено к ней in vivo.
Модель генетического оператора
Теория Жакоба и Моно
В 1961 году появилась статья Франсуа Жакоба и Жака Моно озаглавленная «Генетические механизмы регуляции синтеза белка». Предложенная в ней модель регуляции синтеза белка полностью противоречит всем основным заключениям, к которым мы пришли в гл. I. Эта гипотеза в своей простейшей форме предполагает, что регуляторные механизмы действуют не в цитоплазме клетки, где происходит синтез белка (модель цитоплазматического оператора), а непосредственно на уровне генов, контролируя процесс транскрипции ДНК (модель генетического оператора). Данные, которые Жакоб и Моно положили в основу своей теории, были получены на бактериях и ограничивались по преимуществу их собственными исследованиями регуляции синтеза индуцибельного фермента в-галактозидазы у Е. coli. При обсуждении механизмов действия гена нельзя не остановиться на блестящей статье Жакоба и Моно, и не только потому, что она представляет собой величайший интеллектуальный взлет, но также и потому, что выводы, сделанные в этой статье, противоречат всей совокупности фактов, четко установленных при изучении клеток высших организмов. Если модель генетического оператора, предложенная Жакобом и Моно, верна для бактерий, а модель цитоплазматического оператора — для клеток высших организмов, то приходится признать, что механизмы регуляции синтеза белка в бактериальных клетках и клетках высших организмов принципиально различны. Такое заключение, если бы оно подтвердилось, стало бы одним из важнейших биологических обобщений.
Актиномицин
С тех пор как стали применять антибиотик актиномицин D, появилась возможность значительно расширить круг работ в этом направлении. При высоких концентрациях ктиномицин D связывается с клеточной ДНК и препятствует считыванию с нее РНК. Поэтому его используют, когда хотят произвести своего рода «физиологическую энуклеацию» (хотя этот термин, пожалуй, излишне упрощает действие антибиотика). Следует особо отметить, что актиномицин D чрезвычайно токсичен и при концентрациях, полностью подавляющих транскрипцию ДНК, вызывает быструю гибель большинства клеток. Так что, если высокие дозы актиномицина D подавляют ту или иную физиологическую функцию или какой-либо синтетический процесс, это еще не значит, что исследуемая функция или исследуемый процесс непосредственно и очень тесно связаны с транскрипцией ДНК. Показано, что актиномицин D вызывает серьезные вторичные эффекты, причем гибель клеток обусловлена многими процессами, причины которых только косвенно связаны с прекращением транскрипции ДНК. По этим же причинам нельзя делать никаких заключений о скорости распада матриц для синтеза какого-либо белка, основываясь лишь на изменении скорости синтеза этого белка, которое вызывает актиномицин D. Синтез белков может замедлиться и прекратиться по причинам, не имеющим ничего общего с гибелью матриц. Это не чисто умозрительное соображение: для одного типа клеток показано, что синтез белка, прекращенный актиномицином D, можно полностью восстановить, добавив в культуру глюкозу. Более того, теперь ясно, что высокие дозы актиномицина D могут вызывать быстрое разрушение всех видов клеточной РНК, поэтому делать какие-либо выводы о естественной продолжительности жизни матриц, исходя из опытов с актиномицином D, крайне рискованно.
Соединения пайкой
При пайке деталей ЛA места соединений нагреваются до температуры плавления припоя, которая должна быть ниже температуры плавления соединяемых материалов. Расплавленный припой при этом растекается по нагретым поверхностям стыка деталей, образуя при охлаждении паяный шов, диффузионно и химически связанный с материалом деталей. Пайка позволяет соединять детали не только из однородных, но и из разнородных материалов. Пайкой соединяются, например, стали со сплавами цветных металлов; металлы с графитом, ферритом, фарфором; керамика с полупроводниками; пластмасса, резина и т. п. Кроме того, можно паять детали с тонкостенными элементами, где применение сварки недопустимо из-за опасности прожога. Пайкой можно изготавливать за один производственный цикл сложные по конфигурации узлы и целые конструкции. Это позволяет рассматривать пайку как групповой метод соединения деталей.
В общем машиностроении и в ЛA объем паяных соединений все более увеличивается. Пайка применяется при изготовлении современных камер сгорания жидкостных реактивных двигателей, лопаток турбин, топливных трубопроводов и других конструкций из тугоплавких металлов, плохо поддающихся сварке.
Категория: Рефераты / Производство
Заклепочные соединения
Общие сведения
Заклепочные соединения относятся к классу неразъемных соединений. Они являются одним из наиболее распространенных и надежных способов соединения деталей в общем машиностроении, судостроении и особенно в конструк
ции летательных аппаратов. Например, в широкофюзеляжном самолете до 75% всех соединений выполняются с помощью заклепок. На самолете ИЛ-86 общее число заклепок достигает почти 1, 5 млн штук.
К преимуществам заклепочных соединений относится возможность применения:
□ для соединения несвариваемых деталей;
□ в ответственных конструкциях, воспринимающих большие вибрационные или повторные нагрузки;
□ в конструкциях, не допускающих сварку из-за коробления или опасности отпуска термообработанных детален.
Кроме того, заклепочные соединения по сравнению со сварными более стабильны, лучше контролируются.
Недостатками заклепочных соединений являются:
□ большая масса соединения;
□ более высокая стоимость изготовления;
□ повышенный расход металла;
□ шум и вибрация в процессе ручной клепки.
Категория: Рефераты / Производство
Воздушные компрессоры
Сжатый воздух широко применяется, например, для пуска дизелей или для очистки механизмов при уходе за ними. Воздух под давлением 2,5 МПа и выше обычно получают в многоступенчатом компрессоре. Воздух в компрессоре сжимается сначала в первой ступени, охлаждается и затем сжимается до более высокого давления во второй ступени, затем снова охлаждается и сжимается в следующей ступени. Наиболее часто применяется двухступенчатый компрессор; один из таких компрессоров показан на рис. 7.1.
При ходе всасывания воздух заполняет цилиндр первой ступени через глушитель, фильтр и всасывающий клапан первой ступени. Всасывающий клапан закрывается, когда поршень будет в н. м. т., после чего начинается сжатие воздуха. Когда давление воздуха достигает значения, заданного для первой ступени, начинается нагнетание воздуха через нагнетательный клапан в холодильник первой ступени. Таким же образом происходит всасывание и сжатие в цилиндре второй ступени, в котором благодаря его меньшему объему достигается более высокое давление. После выхода через нагнетательный клапан второй ступени воздух снова охлаждается и подается в баллон сжатого воздуха.
Компрессор имеет жесткий картер, в котором устанавливают три рамовых подшипника коленчатого вала. Блок цилиндров имеет сменные цилиндровые втулки. К движущимся частям компрессора относятся поршни, шатуны и цельный двухколенный коленчатый вал. Сверху на блок цилиндров устанавливается головка цилиндра первой ступени, а на нее — головка цилиндра второй ступени. В обеих головках помещаются всасывающие и нагнетательные клапаны.
Категория: Рефераты / Производство