Когда клетки животных в течение 10—15 мин инкубируют с радиоактивным предшественником РНК, более чем 90% метки обнаруживается в ядре. Тот факт, что радиоактивные предшественники прежде всего включаются в РНК, отмечен еще в 1948 году Бергстрандом, и с тех пор связь между первоначальным появлением метки в ядре и последующим проникновением ее в цитоплазму стала объектом обширной и сложной литературы. Подобное явление наблюдается и у бактерий. Когда бактерии, имеющие хорошо выраженную ядерную зону, например Е. coli, инкубируют в течение 1 мин с радиоактивным предшественником РНК, большая часть метки оказывается в «ядре», а при более длительной экспозиции метка появляется и в цитоплазме бактериальной клетки. В животных клетках быстрометящаяся ядерная РНК находится не в «растворе», а, по-видимому, присоединена к хромосомам. Способ присоединения новообразованной РНК пока еще не известен: возможно, что около ДНК-матрицы ее удерживают водородные связи, однако есть основания предполагать, что белок также принимает в этом участие. Присоединена ли быстрометящаяся РНК к хромосомам бактерий, пока еще не известно. Для того чтобы выделить эту РНК, клетки обычно разрушают и полученный препарат обрабатывают ДНКазой с целью уменьшения его вязкости. Такая обработка, конечно, разрушает бактериальную хромосому и поэтому лишает смысла любое рассуждение о том, было или не было что-либо присоединено к ней in vivo.
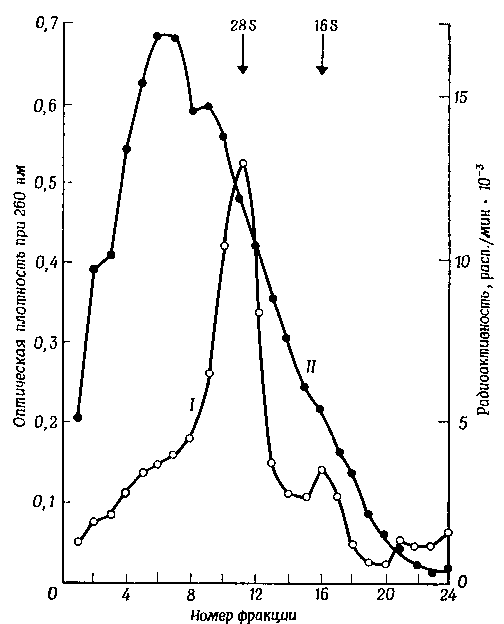
Фиг. 1. Седиментация в градиенте сахарозы ядерной РНК, выделенной из клеток HeLa, после 15-минутной инкубации клеток с 3Н-уридином.
Физические свойства быстрометящейся РНК, выделенной из ядер животных клеток, несколько необычны. Результаты исследования ее в сахарозном градиенте чрезвычайно разнородны. Среди прочих описаны следующие картины седиментации: гетерогенный материал, осаждающийся в той же области, что и 28S- и 163-компоненты рибосом 1 [37]; гетерогенный материал, осаждающийся отчасти вместе с 28S- и 168-компонентами и отчасти несколько медленнее; гетерогенный материал, осаждающийся как в области 28S- и 163-компонентов, так и быстрее ; два широких, но сравнительно гомогенных компонента, выявляемых в области 45 S и 35 S ; в высшей степени полидисперсный материал с коэффициентами седиментации от 80 S до 10 S. Обычно наблюдаемая картина представлена на фиг. 5. Примерно так же обстоит дело и у бактерий. В первых работах Бреннера и др., а также Гро и др. сообщалось, что быстрометящаяся РНК Е. coli дает при осаждении широкий пик с коэффициентом седиментации от 8 S до 16 S, однако позднее в этой РНК была обнаружена большая гетерогенность. Данные, свидетельствующие о том, что эта гетерогенность в основном обусловлена агрегацией молекул быстрометящейся РНК, уже обсуждались ранее.
Недавно более подробно исследовали седиментационные свойства быстрометящейся РНК в животных клетках. Полученные данные также наводят на мысль, что наблюдаемая удивительная полидисперсность — это всего лишь артефакт. Вследствие того что быстрометящаяся РНК почти не обладает вторичной структурой, форма ее молекул и их способность взаимодействовать друг с другом и с иными макромолекулами во многом зависят от метода выделения, концентрации катионов и ионной силы раствора, в котором эта РНК исследуется. Изменяя соответствующим образом физические условия, можно получить почти любой коэффициент седиментации. Однако, когда хорошо очищенный препарат быстрометящейся РНК, отмытый от двухвалентных катионов, исследуют в растворе с низкой ионной силой, то в градиенте сахарозы такая РНК ведет себя как 168-компонент рибосомной РНК.
При осаждении быстрометящейся РНК в присутствии сульфолана, уменьшающего взаимодействие между молекулами РНК, полидисперсность исчезает и при определенных условиях быстрометящийся материал обнаруживается в области меньшего компонента рибосомной РНК . Это очень напоминает наблюдения, сделанные Артманом и др., согласно которым экстрагированная фенолом при низкой ионной силе раствора быстрометящаяся РНК Е. coli дает такую же картину седиментации, как и рибосомная РНК. Но одинаковое поведение веществ в градиенте плотности сахарозы еще совсем не означает, что эти вещества имеют одинаковую молекулярную массу, поэтому для измерения длины полинуклеотидной цепи быстрометящейся РНК применили два других метода. Один из них — переваривание быстрометящейся РНК очищенной экзонуклеазой при определенных условиях, и второй — щелочной гидролиз, позволяющий определить число концевых нуклеотидов по отношению к числу всех нуклеотидов в исследуемых молекулах РНК
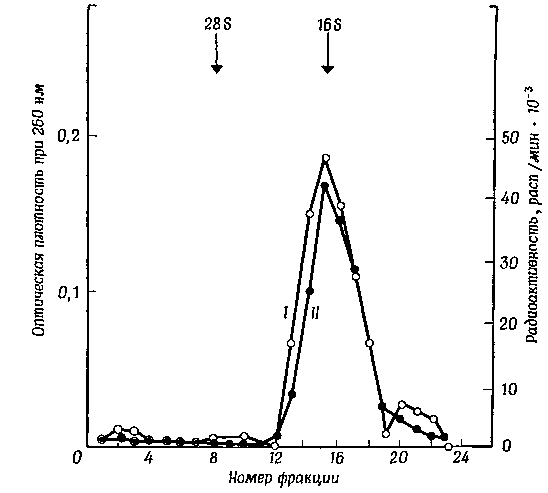
Фиг. 2 Седиментация в градиенте сахарозы ядерной РНК, выделенной из клеток HeLa, после 15-минутной инкубации с 3Н-уридином.
Оказалось, что средняя длина цепи быстрометящейся РНК, определенная обоими методами, очень близка к длине молекулы рибосомной РНК. В правильности этого вывода усомнились Гранбулан и Шеррер, исследовавшие под электронным микроскопом фракции РНК, выделенные с помощью центрифугирования в градиенте плотности. Эти авторы считают, что полидиеперсный материал, осаждающийся быстрее рибосомной РНК, состоит из единичных полинуклеотидных цепей. Однако есть основания усомниться в том, что на электронных микрофотографиях можно отличить единичные полинуклеотидные нити от нитей, которые состоят из двух или более агрегировавших молекул. В самом деле, данные, представленные Гранбуланом и Шеррером, достаточно убедительно показывают, что их препараты содержали какого-то рода агрегаты, поскольку проведенные тем же способом измерения относительно гомогенных компонентов рибосомной РНК выявили значительные различия в длине нитей, причем размер некоторых из них оказался в три раза больше, чем следовало ожидать, исходя из известной молекулярной массы компонентов рибосомной РНК. Следовательно, вряд ли можно надеяться, что измерения, сделанные с помощью электронного микроскопа, дадут нам реальные представления о длине полинуклеотидов.
Тогда, быть может, быстрометящаяся РНК служит просто предшественником рибосомной РНК, как думали раньше? Необычное поведение этой РНК при седиментации еще не доказывает, что она представляет собой рибосомную РНК. Есть основания предполагать, что новообразованная РНК в момент экстрагирования еще прикреплена к хромосоме и не обладает поэтому достаточно жесткой структурой. Большую стабильность, характерную для рибосомной РНК, она могла бы приобрести благодаря метилированию сахара, вторичной модификации некоторых оснований, присоединению белка или же всему этому одновременно. Вопрос о взаимосвязи быстрометящейся и рибосомной РНК можно решить двумя путями: проанализировав кинетику перехода радиоактивной метки во все типы клеточной РНК или определив состав оснований быстрометящейся РНК. Вскоре после того, как обнаружили, что РНК клеточного ядра первая метится радиоактивными предшественниками, было показано, что при переносе клеток, меченных в течение очень небольшого промежутка времени, в нерадиоактивную среду метка исчезает из ядерной РНК и появляется в цитоплазматической РНК. Такие опыты, проводившиеся с использованием как химических, так и радиоавтографических методов, были поставлены на клетках разных видов, в результате чего был сделан вывод, что быстрометящаяся ядерная РНК — это предшественник медленнометящейся цитоплазматической РНК. Подобные опыты, поставленные после того, как были определены седиментационные характеристики рибосомной РНК, показали, что метка, исчезающая из быстрометящейся РНК, появляется в рибосомной РНК, и это послужило причиной считать быстрометящуюся РНК предшественником рибосомной РНК.
Впервые в этом усомнились в 1959 году, когда заметили большое несоответствие между количеством метки, исчезающей из ядерной РНК, и количеством метки, появляющейся в цитоплазматической РНК. В результате многочисленных экспериментов в конце концов установили, что основная часть быстрометящейся ядерной РНК вовлечена в процесс интенсивного внутриядерпого обмена: эта РНК не только быстро синтезируется, но и быстро распадается. Фактически такой вывод был сделан за два года до того, как была высказана гипотеза «посредника», и, следовательно, появился на свет слишком рано, почему и был встречен без особого энтузиазма; однако теперь он подтвержден многочисленными исследованиями, проведенными на самых разных объектах, и сейчас, по-видимому, в основном принят. В клетках, растущих экспоненциально, рибосомная РНК кажется вполне стабильной, причем в неделящихся клетках она заменяется гораздо медленнее, чем быстрометящаяся РНК. Таким образом, независимо от того, является ли быстрометящаяся РНК макромолекулярным предшественником рибосомной РНК или нет, мы можем считать, что на самом деле при обычных экспериментальных условиях большая ее часть не превращается в рибосомную РНК.
Определение состава оснований быстрометящейся РНК связано с огромными методическими трудностями. До самого последнего времени не удавалось ни выделить ее, ни тем более непосредственно определить ее состав с помощью прямого химического анализа. Поэтому широкое применение нашел косвенный метод, предложенный Волкином и Астраханом в 1956 году: в течение короткого промежутка времени клетки инкубировали с радиоактивным фосфатом, а затем исследовали распределение метки в нуклеотидах, полученных после щелочного гидролиза меченой РНК. Считалось, что таким способом можно выяснить состав быстрометящейся РНК. Понятно, что этот метод пригоден для определения нуклеотидного состава только в том случае, если допустить, что радиоактивный фосфор в полинуклеотидах распределяется случайно. Я думаю, достаточно сказать здесь, основываясь на обширных исследованиях, что такое допущение делать нельзя, если клетки инкубируют с радиоактивным предшественником в течение очень небольшого промежутка времени, и поэтому «импульсный фосфорный метод» (как его обычно называют) не годится для определения истинного состава быстрометящейся РНК. Чрезвычайная вариабельность результатов, полученных этим методом, открытие фракций РНК, близких к ДНК по составу оснований, равно как и частые неудачи при выделении таких ДНК-подобных фракций, а также резкие колебания состава быстро метящейся РНК, вызванные внешними стимулами или изменением культуральных условий, — все это означает, что распределение метки в РНК после импульсного введения радиоактивного фосфора обусловлено на самом деле не составом синтезированной за это время РНК, а совсем другими причинами.
Основываясь на необычных физических свойствах быстрометящейся РНК, удалось разработать метод ее выделения, а следовательно, и прямого определения ее состава. Возможно, что именно вследствие недостаточной выраженности вторичной структуры быстрометящаяся РНК хорошо задерживается на кизельгуровых колонках с метилированным альбумином, если использовать такие концентрации солей, при которых элюируются все другие рибонуклеиновые кислоты клетки.
РНК, задерживающаяся на колонках с метилированным альбумином, обычно содержится в клетках в ничтожных количествах, но недавно обнаружили, что она накапливается в ядрах при резких шоковых воздействиях на клетку. Из таких клеток легко выделить достаточное количество быстрометящейся полидисперсной РНК, чтобы определить в ней после щелочного гидролиза содержание всех четырех нуклеотидов.
Оказалось, что состав оснований быстрометящейся РНК соответствует составу 16S-PHK, выделяемой из меньшей субъединицы рибосом. Полученные результаты подтверждают точку зрения, согласно которой большая часть быстрометящейся РНК представляет собой предшественник по крайней мере одного из компонентов рибосомной РНК. Такое представление позже подтвердилось также тем, что распределение метилированных остатков рибозы между последовательностями оснований в быстрометящейся РНК не отличается от распределения их в рибосомной РНК. Если быстрометящаяся РНК действительно служит предшественником рибосомной 16S-PHK, то ее быстрый обмен может означать, что при обычных экспериментальных условиях лишь очень небольшая часть предшественника превращается в зрелую рибосомную РНК. Несколько позже я подробнее остановлюсь на этом вопросе.
PHK
При определенных условиях экстрагирования и центрифугирования часть быстрометящейся РНК может осаждаться в виде довольно широкого пика в той области сахарозного градиента, которая приблизительно соответствует коэффициенту седиментации 45 S. Когда клетки после короткой экспозиции с радиоактивным урацилом переносят в нерадиоактивную среду, метка покидает область 45S-РНК и появляется в области рибосомной 28S- и 16S-PHK; такой же процесс, но несколько менее выраженный, продолжается и в присутствии умеренных доз актиномицина D. Это навело на мысль, что 45S-PHK служит предшественником рибосомных 28S- и 16S-PHK. А поскольку5S-PHK представляет собой единую ковалентно связанную полинуклеотидную цепь, предположили, что рибосомные 28S- и 16S-PHK возникают при асимметричном расщеплении этой единой гигантской макромолекулы предшественника.
Но так как этот предшественник (исходя из простого соотношения длины цепи и коэффициента седиментации) должен быть примерно в два раза длиннее молекулы, составленной из одной 28S- и одной 168-субъединицы, пришлось далее допустить, что около половины исходной молекулы разрушается при отщеплении 28S и 16Э-компонентов.
С первого же взгляда видно, что по существу своему эта схема неправдоподобна, и потребовались бы очень веские доказательства, чтобы принять ее. На самом деле результаты работ, проведенных в последние годы, сделали ее еще менее приемлемой. Прежде всего, если 28S- и 16S-компоненты образуются при разделении единой большой молекулы, то следует ожидать, что при инкубации клеток с радиоактивным уридином метка должна равномерно включаться в 28S- и 16S-PHK. Однако сначала она появляется в ядерной 16S-PHK и только потом в ядерной 28S-PHK, и в дальнейшем характер распределения метки в этих двух компонентах легко изменять с помощью различных процедур.
Две группы исследователей изучали распределение метки в олигонуклеотидах, полученных после частичного гидролиза 16S- и 28S-PHK, которые выделяли из клеток, импульсно-меченных радиоактивным уридином. Обе группы пришли к выводу, что включение метки в 28S- и 16S-РНК происходит по-разному. Этот результат трудно совместить с предположением о том, что 28S- и 168-компоненты возникают при разделении одного и того же полинуклеотида Более того, Брэмвелл показал, что РНК, неотличимая от 16S-PHK ни по составу оснований, ни по коэффициенту седиментации, может накапливаться в клеточном ядре, когда синтез 28S-PHK полностью подавлен низкими дозами актиномицина D. Все сказанное выше свидетельствует о том, что предположение о возникновении 28S- и 16S-PHK в результате расщепления одного общего гигантского предшественника маловероятно. Такую схему вряд ли предложили, если бы не мысль, что полидисперсные компоненты РНК, осаждающиеся быстрее, чем рибосомная РНК, представляют собой единые ковалентно связанные полинуклеотиды необычной длины. Аргументы против этого предположения были приведены ранее.
Используемая литература: Г. Харрис
Перевод с английского: М. И. Маршак
Ядро и цитоплазма: под ред. и с предисловием д-ра биол. наук Н. И. Шапиро
Москва 1973 год.
Скачать реферат:
Пароль на архив: privetstudent.com