НАРУЖНАЯ ПОВЕРХНОСТЬ спинного МОЗГА И ВНУТРЕННЕЕ РАЗДЕЛЕНИЕ
Спинной мозг с дорсальной стороны имеет дорсальный срединный желоб, sulcus medianus dorsalis, и по бокам латеральные дорсальные желоба, sulcus lateralis dorsalis. По последним волокна корешков входят дорсально в спинной мозг. Соответствующие вентролатерально расположенные желоба для выхода вентральных корешков практически не выражены. С дорсальной стороны спинной мозг разделён дорсальной срединной перегородкой, septum medianum dorsale, с вентральной стороны имеется глубокая, заполненная pia mater (соединительной тканью и сосудами) вентральная срединная щель, fissura mediana ventralis.
В спинном мозге по периферии серого вещества, substantia grisea, располагающегося в виде буквы Н, находится белое вещество, substantia alba, в котором проходят волокна восходящих и нисходящих путей.
Строение спинного мозга на поперечном сечении различается у разных видов животных, а также зависит от сегмента, в котором находится. Различия наблюдаются и в соотношении серого и белого вещества.
Серое вещество спинного мозга, располагающееся в дорсальном и вентральном рогах, cornu dorsale etcomu ventrale, разделяется находящимися между этими областями латеральным промежуточным веществом, substantia intermedia lateralis, а также серой спайкой, comissura grisea (центральное промежуточное вещество, substantia intermedia centralis). Посередине спайки расположена оставшаяся часть обширного эмбрионального пространства (нервной трубки), которое в виде центрального канала, canalis centralis, проходит по всей длине спинного мозга. Форма центрального канала чаще всего не округлая, а в виде удлинённого овала, причём высота его у кошек составляет около 100 μm, а ширина — около 50 μт. У собак, согласно Biach (1906), значения сходные. Центральный канал разделяет серую спайку на comissura grisea dorsalis и comissura grisea ventralis.
Хотя название "рог” соответствует строению структур на поперечном сечении спинного мозга и допускается в виде анатомического названия, следует иметь в виду, что речь идёт о столбах (columnae), также как белое вещество состоит из отдельных пучков, номенклатуру которых следует принимать во внимание.
Организация серого вещества
Согласно иерархическому принципу строения центральной нервной системы, в ней располагаются центры, которые самостоятельно и независимо управляют процессами, происходящими на периферии (тонус, рефлексы). Это значит, что в substantia grisea располагаются важные переключательные центры. Кроме того, здесь лежат центры вегетативной нервной системы. Морфологически это выражается в формировании ядер, nuclei; они представляют собой группы клеток, различные по форме и размеру.
Без учёта сегментных различий в верхней части дорсального рога располагается желатинозное вещество, substantia gelatinosa, и краевая зона, zona marginalis (-/f). В дорсальном роге находятся собственное дорсальное ядро, nucleus dorsalis proprius и грудное ядро, nucleus thoracicus (-/f). В средней части находятся вегетативные ядра (симпатические, парасимпатические): промежуточное латеральное ядро, nucleus intermediolateralis, и промежуточное медиальное ядро, nucleus intermediomedialis, в вентральном роге двигательное ядро, nucleus motorius, с клетками соматомоторных волокон (-/к). Кроме этих описанных групп клеток имеются нейроны-переключатели, клетки спаек и ассоциативные клетки (-/m, n, о), которые имеют соединения. Кана-тиковые клетки (-/р) посылают свои аксоны в путь (tractus), соединяющие спинной мозг с головным мозгом.
Хотя нейроны серого вещества спинного мозга изучаются уже более сотни лет, и принципы их объединения в ядра известны, их локализация с обычными топографическими и функциональными обозначениями (дорсальный рог, промежуточное серое ядро, двигательное ядро и т. д. ) несовершенна и может допускать неправильное толкование. При использовании окраски по Нисслю, на 100 μm разрезе спинного мозга кошки Rexed (1952, 1954) расположил тела нейронов по форме, размеру, плотности и их распределению, а в организации серого вещества выявил 9 пластин (laminae). Пластины, в основном, проходят через весь спинной мозг. В дорсальном роге они наиболее развиты, а вентрально, прежде всего в области утолщений выражены менее из-за значительной выраженности мотонейронов. Кроме 9 пластин (I-XI) Rexed обозначил оставшееся центральное промежуточное вещество (серое) цифрой X.
Данная классификация не охватывает одно ядро, лежащее вентролатерально от дорсального рога, латеральное шейное ядро, nucleus cervicalis lateralis. У кошек это ядро располагается в С1 и С2 и рострально прилегает к сетчатой формации, formatio reticularis (Rexed / Brodal, 1951). У собак это ядро простирается от задвижки Obex и С3. Ядро содержит кожные афферентные волокна из дорсального рога и по tractus spinocervicalis, который проходит ипсилатерально в funiculus dorsalis.
У кошек ядро организовано соматотоиичес-ки. Задняя конечность представлена дорсолате-рально, передняя конечность вентромедиально (Craig Tapper, 1978; Svenson et al., 1985). Имеется ли у собак такая же соматотопическая организация, в имеющейся литературе остаётся невыясненным.
Tractus spinocervicalis идёт от рецепторов, которые регистрируют небольшие быстрые изменения окружающей среды. Он осуществляет реакцию на лёгкое движение волос или механическое смещение кожи (Oswaldo-Cruz/Kidd, 1964). Nucleus cervicalis — добавочное ядро-реле у собак и кошек (у людей это ядро рудиментарное; Kitai et al., 1965). Оно включается в tractus spinocervico-thalamicus, путь, который присоединяется к медиальной петле, lemniscus medialis, к противоположной стороне и заканчивается в nucleus ventralis caudolateralis таламуса (Boivie, 1970; Hagg/Ha, 1970).
В шейном отделе спинного мозга серое вещество между дорсальным и вентральным рогом образует сетчатую (ретикулярную) формацию, formatio reticularis.
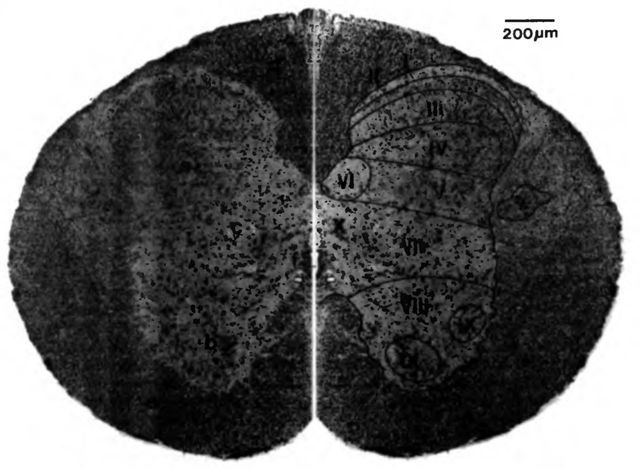
Рис. 6. Поперечный разрез спинного мозга (1/2 шейный сегмент) кошки 2-дневного возраста, трёхцнетная окраска (по Seiferle, 1992)
Левая половина: a cornu dorsale; b cornu ventrale; с substantia intermedia, d funiculus dorsalis; e funiculus ventrolateralis; f canalis centralis
Правая половина: laminae (I - IX) серого вещества, в примерном расположении, по Rexed (1952, 1954).
Также вегетативная нервная система имеет центры в спинном мозге. Симпатическая часть нервной системы формируется в области латерального промежуточного вещества, substantia intermedia lateralis грудной и поясничной частей спинного мозга, спинномозговой центр парасимпатического отдела нервной системы локализуется в крестцовом отделе спинного мозга. У собак это прерывный столб клеток (промежуточное латеральное ядро, nucleus inter-mediolateralis) от каудальной трети S1 до средней трети S3. Основное скопление клеток располагается в средней трети S2 (Samson/Reddy, 1983).
Организация белого вещества
Белое вещество каждой стороны подразделяется в зависимости от расположения волокон корешков на дорсальный канатик, funiculus dorsalis (между дорсальным корешком и дорсальной перегородкой), латеральный канатик, funiculus lateralis (между дорсальным и вентральным корешком) , а также вентральный канатик, funiculus ventralis (между вентральным корешком и fissura mediana ventralis) (-/III). Располагающееся между дорсальными рогами белое вещество полностью разделяется на две половины срединной дорсальной перегородкой, septum medianum dorsale (-/ D). Оба вентральных канатика связывает белая спайка, comissura alba (ventralis), располагающаяся вентраль-но от серой спайки (-/3). С дорсальной стороны соответствующая структура отсутствует.
Такое разделение белого вещества — а оно на поперечном сечении спинного мозга представлено, преимущественно, поперечными сечениями аксонов — не даёт представления об отличиях функционально различных проводящих путей друг от друга. У домашних млекопитающих эти пути не так изучены, как у людей. Для этого следует учитывать сегментарные различия. Они особенно касаются дорсального канатика. Эта область формируется из восходящих волокон, которые проводят импульсы чувствительности (прикосновения и чувствительности к надавливанию) без переключения в спинном мозге от периферии к продолговатому мозгу, medulla oblongata. Волокна из задней части тела, особенно от задней конечности, формируют тонкий пучок, fasciculus gracilis (-/7), который по срединной линии примыкает к septum medianum dorsale, в то время как волокна из передней части тела, особенно из передних конечностей, присоединяются латераль-но, формируют клиновидный пучок, fasciculus cuneatus (-/8) и сначала появляются в грудном и шейном отделах спинного мозга. Оба пучка по направлению к голове на дорсальной стороне спинного мозга заметны как тяжи, которые разделяются срединным дорсальным желобом, sulcus intermedius dorsalis. Из-за соединения спинной мозг — продолговатый мозг (Bulbus medullae spinalis) оба канатика объединяются в tractus spinobulbaris.
Таблица 1. Локализация двигательных нейронов в спинном мозге кошки, установлена методами дегенерации (DEG) или, соответственно, ретроградной маркировкой (HRP). Если нервных стволов не имеется, в скобках указаны мышцы, в которые инъецируются маркирующие вещества.
|
Сегмент |
Нерв (мышца) |
Автор |
|
С1-С4 |
шейные нервы (m. splenius, m. biventer cervicis, m. complexus) |
Richmond et al., 1978 (HRP) |
|
С2-С6 |
ramus externus n. accessorii (m. trapezius) |
Vanner/rose, 1984 (HRP) |
|
С5 —Th1 |
ramus profundus n. radialis |
Fritz et al., 1981 (HRP) |
|
С6 —Th1 |
n. radialis |
Fritz et al., 1986 a (HRP) |
|
С6—Thl |
n. medianus (m. flexor carpi radialis) |
Iwamoto et al., 1980 (HRP) |
|
С6-С8 и Th9 — L3 |
вентральные ветви (m. latissimus dorsi) |
Holstege et al., 1987 (HRP) |
|
С7-Тh1 |
n. medianus и n. ulnaris |
Fritz et al., 1986 b (HRP) |
|
С7/8- Th1 |
n. ulnaris (m. flexor carpi ulnaris) |
Iwamoto et al., 1980 (HRP) |
|
С8 —Th1 |
вентральные ветви (m. cutaneus trunci) |
Holstege et al., 1987 (HRP) |
|
C8-L2 |
вентральные ветви (мышцы живота) |
Holstege et al., 1987 (HRP) |
|
Th1 — L4 |
вентральные ветви (m. longissimus dorsi) |
Holstege et al., 1987 (HRP) |
|
Th4 — L3 |
вентральные ветви (мышцы живота) |
Miller. 1987 (HRP) |
|
L4-S1 |
двигательные нервы задней конечности |
Romanes, 1951 (DEG) |
|
L6-L7 |
n. fibularis (m. tibialis cranialis) |
Iliya/dum, 1984 (HRP) |
|
L6-S1 |
n. tibialis (m. gastrocnemius) |
Weeks/English, 1985 (HRP) |
|
L6-S1 |
n. fibularis (m. fibularis longus, m. fibularis brevis, m. fibularis tertius) |
Horcholle-Bossavit et al., 1988 (HRP) |
|
L6-S1 |
n. fibularis |
Balthasar, 1952 (DEG) |
|
L7 — S1 |
n. tibialis (m. gastrocnemius, m. soleus) |
Burke et al., 1977 (HRP) |
|
L7-S2 |
n. tibialis |
Balthasar, 1952 (DEG) |
|
S2 —S3 |
nn. coccygei et nn. sacrales (хвостовые мышцы) |
Wada et al., 1990 (HRP) |
Короткие нисходящие ветви названных пучков прилегают к перегородке или формируют в шейном отделе спинного мозга каплеобразную область между тонким и клиновидным пучками, запятую Шультца.
Другим восходящим путём, который проводит чувствительные импульсы, является tractus spinothalamicus. Он проходит у кошек дорсолатерально. Этот проводящий путь возникает практически только в lamina I, все нейроны которой без исключения проводят импульсы болевой чувствительности (Jones et al., 1985). Этот проводящий путь отличается от такого у человека (Kennard, 1954). Он заканчивается ипсилатерально в различных ядрах промежуточного мозга, однако не в nucleus ventralis caudolateralis, как у человека. Соматотопи-ческая организация нервных окончаний у кошек также не проявляется (Boivie, 1971).
Вентролатеральный канатик не отграничивается от внешней среды. Хотя у домашних млекопитающих дифференциация белого вещества невозможна в полной мере, как у людей, и поэтому в строении спинного мозга присутствуют "белые пятна”, отличия от строения спинного мозга у человека имеются, и они могут быть описаны только вкратце. Латеральный кортикоспинальный путь, tractus corticospinalis lateralis, по сравнению с таковым у человека, относительно небольшой, однако у собак и кошек выражен лучше, чем у других видов домашних животных. Tractus corticospinalis лежит вентрально на medulla oblongata, и из-за своей формы у людей назван пирамидным путём. Он возникает вgyri sygmoideus, coronarius et ectosylvius lateralis и заканчивается у плотоядных в крестцовом отделе спинного мозга. Его основная часть идёт в виде перекрещенного пути tractus corticospinalis lateralis, в то время как небольшой неперекрещенный вентральный кортикоспинальный путь, tractus corticospinalis ventralis, ограничен верхними шейными сегментами (Chamber/Liu, 1957). Наличие у домашних млекопитающих tractus corticospinalis ventralis оспаривается. Tractus corticospinalis у кошек заканчивается в laminae IV-VII спинного мозга (Nyberg-Hansen/Brodal, 1963). У собак имеется ипси- и контралатеральные нервные окончания в laminae V, VI, VII et VIII, то есть волокна tractus corticospinalis заканчиваются интернейронами, и только небольшая часть — в лежащих в lamina IX двигательных нейронах. У кошек имеется ещё незначительная часть от прямого переключения волокон пирамидного пути на двигательные нейроны (Buxton, 1967).
У кошек было насчитано 415000 аксонов в tractus corticospinalis, 88% которых составляли мие-линизированные, а 12% — немиелинизированные волокна (Biedenbach et al., 1986). У собак, как и у людей, 55% волокон заканчивается в шейном отделе спинного мозга, 20% — в грудном отделе и 25% в поясничном и крестцовом отделах спинного мозга (Lassek et al., 1930). Lassek/Rasmunssen (1940) насчитали у кошек 186000, а у собак 285000 волокон в tractus corticospinalis.
Эта пирамидно-двигательная система играет у домашних млекопитающих подчинённую роль. Пересечение пирамидного пути у собак и кошек около medulla oblongata незначительно влияет на двигательные функции. Наибольшие изменения возникают при повреждении двигательной коры. Из этого делается вывод о влиянии двигательной коры на экстрапирамидальную моторику (Marshall, 1936): имеются только небольшие соединения cortex cerebri с двигательными нейронами в спинном мозге. На первом месте для обеспечения моторики у домашних млекопитающих находится вентрально примыкающий tractus reticulospinalis lateralis и rubrospinalis (-/16).
Красноядерно-спинномозговой путь, tractus rubrospinalis у кошек перекрещен. Волокна, заканчивающиеся в шейном отделе спинного мозга, выходят из дорсальных и дорсомедиальных частей красного ядра, nucleus ruber, волокна для поясничного отдела спинного мозга — из вентральных и вентролатеральных частей (Pompeiano/Brodal, 1957). Соматотопическая организация tractus rubrospinalis была подтверждена (Nyberg-Hansen/ Brodal, 1964). Было также показано, что Nucleus ruber проецируется непосредственно на двигательные нейроны в С8 и Th1 (Holstege, 1987) или, соответственно, на ядра нервов для мускулатуры пальцев (McCurdy et al., 1987).
Ретикулоспинальный путь, tractus reticulospinalis, идёт от моста и продолговатого мозга. Волокна моста идут с той же стороны в funiculus lateralis. Они многочисленны, как медуллярные волокна, и заканчиваются в laminar VIII VII. Медуллярные волокна идут билатерально вентрально в funiculus lateralis и заканчиваются, прежде всего, в lamina VII vNvberg-llansen, 1965).
Вестибулоспинальный путь, tractus vestibulospi nаIis, проходит сначала в funiculus ventrolateralis, затем отклоняется дорсомедиально и идёт в поясничном отделе спинного мозга к fissura mediana ventralis. Волокна заканчиваются в lamina VIII, не на моторных нейронах, а в nucleus thoracicus или на нейронах латерального промежуточного вещества (Nvberg-Hansen Mascitti, 1964).
Покрышечно-спинномозговой путь, tractus tectospineilis, у кошек полностью перекрещивается и заканчивается в С1-С4 в laminae VII, VI и VIII. Этот путь не имеет прямых соединений с моторными нейронами (Nyberg-Hansen, 1964).
Восходящие и нисходящие проводящие пути не прилегают непосредственно к серому веществу. Узкая полоска белого вещества соединяет в виде основных пучков (fasciculi proprius ventrolateralis et proprius dorsalis) восходящие и нисходящие сегменты (ассоциативные клетки) или правую и левую стороны (комиссуральные клетки). Основные пучки являются самостоятельными органами регуляции спинного мозга (рефлекторные органы) и формируют вместе с функционирующим сходным образом дорсолатеральным трактом, tractus dorsolateralis (zona terminalis), занятой Шульца (-/5) и медиальным продольным пучком (fasciculus longitudinalis medialis) (-/4), а также переключательными, комиссуральными и ассоциативными нейронами в сером веществе собственный аппарат спинного мозга, который противопоставляется проводниковому аппарату (спинной мозг как проводящий орган между высшими центрами, головным мозгом и периферией).
В целом, функциональная связь отражается на поперечном сечении спинного мозга.
Используемая литература: Анатомия собаки и кошки (Колл, авторов) / Пер. с нем. Е. Болдырева, И. Кравец. - М.: «АКВАРИУМ БУК», 2003. 580с., ил. цв. вкл.
Скачать реферат: