Головной мозг крысы
Головной мозг — encephalon — является передним отделом центральной нервной системы, расположенным в полости черепа.
В период эмбрионального развития после замыкания нервной пластинки в трубку в её переднем отделе образуются мозговые пузыри — vesiculae cerebrales — расширения нервной трубки, пред ставляющие зачатки отделов головного мозга.
Различают три мозговых пузыря: первичный задний мозг, средний мозг и первичный передний мозг. В течение эмбрионального Развития передний и задний мозговые пузыри делятся каждый на две части и, таким образом, возникают 5 мозговых пузырей, преобразующихся в ходе развития в пять отделов головного мозга.
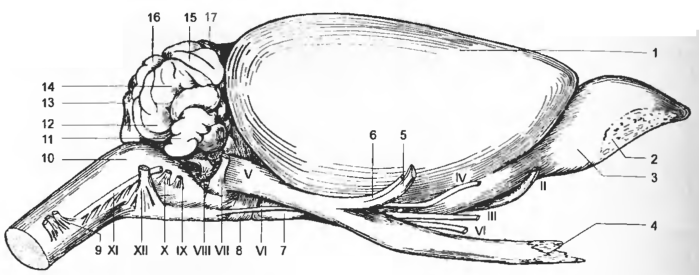
Рис. 1 Мозг с латеральной поверхности
1 — hemispherium, 2 — regio nn. olfactorii, 3 — bulbus olfactorius, 4 — n. ophthalmicus, 5 - n. mandibularis, 6 — n. maxillaris, 7 — hypophysis, 8 — pons, 9 — СI, 10 — medulla oblongata, 11— parafloeculus dors., 12 — lobulus paramedianus, 13 — lobus caud., 14 — lobulus ansiformis, 15 — lobulus simplex, 16 — pyramis vermis, 17 — lobus rostralis; II-XII — nn. craniales.

Рис. 2 Отделы головного мозга и расположенные в них основные структуры
Из заднего мозгового пузыря развивается ромбовидный мозг — rhombencephalon, состоящий из продолговатого мозга — myelencephalon - и заднего мозга — metencephalon (рис. 10.8). Из среднего мозгового пузыря образуется средний мозг — mesencephalon. Для того чтобы отразить представление «ствол мозга», введен новый термин мозговой ствол — truncus encephalicus, включающий продолговатый мозг, задний мозг (мост) и средний мозг. Из переднего мозгового пузыря развивается передний мозг — prosencephalon, состоящий из промежуточного мозга — diencephalon — и конечного мозга — telencephalon. В процессе развития каждый из этих отделов мозга дифференцируется. Миеленцефалон является продолговатым мозгом; задний мозг подразделяется на мозжечок и мост; средний мозг делится на ножки мозга и крышу (среднего мозга); промежуточный мозг состоит из гипоталамуса и таламуса с окружающими образованиями; конечный мозг дает начало большому и обонятельному мозгу и полосатому телу. Полости мозговых пузырей преобразуются в полости головного мозга — желудочки мозга. Полость ромбовидного мозга превращается в IV желудочек, который сообщается с центральным каналом спинного мозга. В среднем мозгу полость мозгового пузыря образует водопровод среднего мозгу, в промежуточном — III желудочек, а в конечном мозгу — боковые желудочки.
Продолговатый мозг
Продолговатый мозг (бульбус) — medulla oblongata (bulbus, myelencephalon) — часть мозгового ствола, протянувшаяся от первой пары шейных нервов до моста. Каудально продолговатый мозг без отчетливой границы переходит в спинной мозг. Продолговатый мозг вместе с задним мозгом составляет ромбовидный мозг. На вентральной поверхности продолговатого мозга хорошо заметна срединная (вентральная) щель — fiss. mediana (ventr.), являющаяся продолжением одноименной щели спинного мозга. По сторонам срединной щели проходит слабо выраженная парная вентральная латеральная борозда — sul. ventr. lat. — место выхода отводящего нерва (ростральный участок борозды) и подъязычного нерва (каудальный участок). По сторонам от срединной щели располагается парное продольное возвышение — пирамида (продолговатого мозга) — pyramis (medullae oblongatae), содержащая волокна пирамидного пути. Каудально пирамиды сужается и образуют перекрест пирамид (двигательный перекрест) — decussatio pyramidum. Латерально от пирамиды находится небольшое несколько вогнутое парное образование треугольной формы — олива — oliva. Латеральнее и ростральнее места выхода подъязычного нерва отходят добавочный, блуждающий и языкоглоточный нервы. На дорсальной поверхности продолговатого мозга проходит срединная (дорсальная) борозда — sul. medianus (dors.), продолжающая срединную борозду спинного мозга рострально к IV желудочку до его задвижки. Срединная борозда разделяет продолжения дорсальных канатиков спинного мозга. Латеральнее срединной борозды проходит слабо заметная парная дорсальная латеральная борозда — sul. lat. dors., идущая вдоль дорсальной стороны продолговатого мозга по направлению к латеральному карману (IV желудочка). Борозда с каждой стороны отделяет дорсальный канатик от продолжения латерального канатика спинного мозга. Расположенная медиальнее дорсальной латеральной борозды, едва заметная дорсальная промежуточная борозда — sul. intermedius dors. — отделяет тонкий пучок дорсального канатика от клиновидного пучка, которые являются продолжениями одноименных пучков спинного мозга. Тонкий пучок оканчивается бугорком тонкого ядра — tub. nuclei gracilis — небольшим возвышением на уровне тонкого ядра. Ростральный конец клиновидного пучка несколько расширяется и образует возвышение на уровне клиновидного ядра бугорок клиновидного ядра — tub. nuclei cuneati.
В продолговатом мозгу представлено значительное количество ядер, пучков и путей. Здесь находятся тонкое ядро — nucl. gracilis. клиновидное ядро — nucl. cuneatus и некоторые ядра черепных нервов: главное чувствительное ядро тройничного нерва — nucl. sensorius principalis n. trigemini, ядро спинномозгового пути тройничного нерва — nucl. tr. spinalis n. trigemini, двигательное ядро подъязычного нерва — nucl. motorius n. hypoglossi и дорсальное двигательное ядро блуждающего нерва — nucl. motorius dors, n. vagi. Кроме того, здесь расположены: нижнее оливное ядро — nucl. olivaris inf., латеральное ретикулярное ядро — nucl. reticularis lat., двойное ядро — nucl. ambiguus, ядро одиночного пути — nucl. tr. solitarii, а также многочисленные вестибулярные — nucll. vestibulares, улитковые — nucll. cochleares, ретикулярные ядра — nucll. reticulares, и ядра шва — nucll. raphe.
Задний мозг
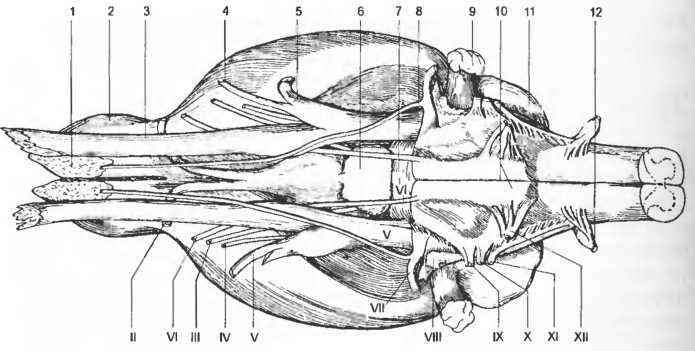
Рис. 3 Мозг с вентральной поверхности
1 — regio nn. olfactorii, 2 — bulbus olfactorius, 3 — n. ophthalmicus, 4 — n. maxillaris, 5 — n. mandibularis, 6 — hypophysis, 7 — pons, 8 — n. petrosus major, 9 — paraflocculus dors., 10 — pyramis (medullae oblongatae), 11 — cerebellum, 12 — CI; II—XII — nn. craniales.
Задний мозг — metencephalon — является ростральной частью ромбовидного мозга, расположенной между продолговатым и средним мозгом; состоит из моста и мозжечка. Здесь же приводится описание IV желудочка.
Мост — pons, часто именуется варолиевым мостом — pons Varolii, является отделом головного мозга, находящимся между продолговатым и средним мозгом. Состоит из поперечных волокон — fibrae pontis transversae — и клеточных групп корково-мозжечковых проводящих путей и ростральной части покрышки ромбовидного мозга. У крысы развит относительно слабо.
Латерально пучки волокон моста соединяются с мозжечком посредством средней мозжечковой ножки (плечика моста) — pedunculus cerebellaris medius (brachium pontis — BNA, JNA), содержащей афферентные мостомозжечковые волокна — fibrae pontocerebellares.
От латеральной части моста двумя корешками — моторным и двигательным отходит тройничный нерв (V). Маленький моторный корешок - rad. motoria n. trigemini — лежит медиальнее тройничного узла большого чувствительного корешка — rad. sensoria n. trigemini.
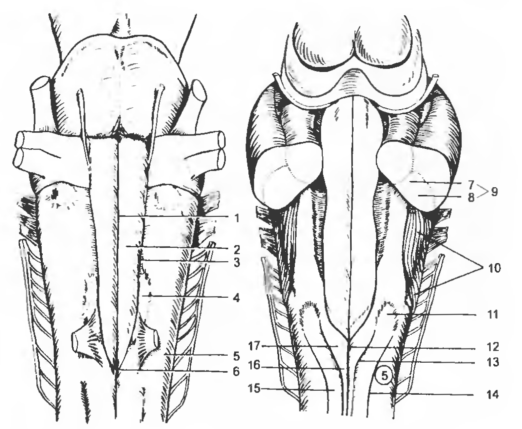
Рис. 4 Продол говатый мозг и мост с вентральной (А) и дорсальной (Б) поверхностей
1 - fiss. mediana (ventr.), 2 - pyramis, 3 — sul. lat. ventr.,4 - oliva, 5 — funiculus lat., 6 — decusatio pyramiduin, 7 — corp. juxtarestiforme, 8 — corp. restiforme, 9 - pedunculus cerebellaris caud., 10 — fibrae arcuatae super -ficialis, 11 — tub. nucl. cuneati, 12 — sul. medianus (dors.), 13 — sul. intermedius dors., 14 — sul. lat. dors., 15— fasc. cuneatus, 16 — fasc. gracilis, 17 — tub. nucl. gracilis.
У границы с продолговатым мозгом находится трапециевидное тело — corp. trapezoideum — плоский пучок поперечно идущих волокон от улитковых ядер.
Мост подразделяется на вентральную базилярную часть — pars basilaris pontis — и дорсальную часть моста, или покрышку моста, — pars dors, pontis s. tegmentum pontis. В мосту расположены многочисленные ядра, волокна и пути. Среди них ядра моста — nucll. pontis, ретикуло покрышечное ядро — nucl. reticulotegmentalis, срединное ядро шва — nucl. raphe medianus, ядро шва моста — nucl. raphe pontis, голубоватое ядро — nucl. ceruleus и некоторые ядра черепных нервов: двигательное ядро тройничного нерва — nucl. motorius n. trigemini и ядро лицевого нерва — nucl. n. facialis. Кроме того, в ретикулярной формации моста представлены множественные структуры: ретикулярное ядро моста — nucl. reticularis pontis, вентральное ядро покрышки — nucl. pedunculopontinus tegmenti, паралемнисковое ядро — nucl. paralemnisci и др. Наконец, в вентральной области моста расположена группа слуховых ядер, объединенных в понятие верхний оливный комплекс — oliva. sup., состоящий из медиальной и латеральной верхних олив, верхнего околооливного ядра — nucl. paraolivaris sup., дорсального и вентрального ядер трапециевидного тела — nucll. dors, et ventr. corporis trapezoidei.
Мозжечок — cerebellum — является частью заднего мозга, расположенной дорсальнее ромбовидной ямки. Состоит из непарной срединной части — червя — vermis — и двух латеральных участков — полушарий мозжечка — hemespherium cerebelli. Поверхность мозжечка покрыта многочисленными тонкими пластинообразными извилинами — листками мозжечка — folia cerebelli, разделенными глубокими и узкими щелями мозжечка — fissurae cerebelli. Борозды между червем и полушариями и между отделами полушарий именуются бороздами мозжечка — sulci cerebelli. Выделяют тело мозжечка — corp. cerebelli, состоящее из ростральной доли — lobus rostralis — и каудальной доли — lobus caud. — и отделенное от клочка и околоклочка околоклочковой бороздой — sul. parafloccularis. Мозжечок соединен с другими частями мозга тремя отчетливыми пучками волокон — ножками мозжечка — pedunculi cerebelli. Каудальная (нижняя) мозжечковая ножка — pedunculus cerebellaris caud. (inf.) соединяет мозжечок с продолговатым мозгомПодразделяется на веревчатое тело — corp. restiforme — основную латеральную часть, содержащую многочисленные пути афферентных волокон, и вблизиверевчатое тело — corp. juxtarestiforme — медиальную, небольшую часть, содержащую преддверно-мозжечковый путь. Ростральная (верхняя) мозжечковая ножка (соединительное плечико) — pedunculus cerebellaris rostralis (sup.) (brachium conjunctivum BNA, JNA) — соединяет мозжечок со средним мозгом.

Рис. 5 Мозжечок (графиче ская реконструкция на основании серийных срезов); справа на рисунках удалена ростральная доля для выявления простои дольки. Вид с каудальной (А), дорсальной (Б), ростральной (В), вентральной (Г) сторон и на срединносагиттальном срезе (Д)
1-10 - lobuli cerebelli, ApmF — fiss. ansoparamediana, Cl — crus 1 (lobulus ansiformis), C2 — crus 2 (lobulus ansiformis), COP — cupula pyramidis, FL — flocculus, IcF — fiss. intercruralis, icp - pedunculus cerebellaris caud., jrb — corp. juxtarestiforme, mcp — pedunculus cerebellaris medius, PFL — paraflocculus, PIF — fiss. posterolat., PflS — sul. parafloccularis, PM — lobulus paramedianus, PpF — fiss. prepyramidalis, PreculF — fiss. preculmeninis, PrF — fiss. prima, PsF — fiss. caud. sup., scp — pedunculus cerebellaris rostralis, SecF — fiss. secunda, Sim — lobulus siniplex, smv — velum medullare rostrale.
Деление мозжечка на доли и дольки, предложенное Ларселлом (Larsell, 1952), используется и в настоящее время; параллельно с этой номенклатурой мы приводим и традиционные термины. Мозжечок подразделяется в ростро-каудальном направлении на 10 долек, обозначаемых цифрами от 1 до 10. Глубокая первая щель — fiss. prima — отделяет ростральную (переднюю) долю — lobus rostralis — от каудальной (задней) доли — lobus caud. Первая щель и предвершинная щель — fiss. preculmeninis, делящие ростральную долю на дольки 1—3 и 4 5, расщепляются на их ростральной и каудальной стенках и достигают латерального края мозжечка. Другие междольковые щели ростральной доли не идут так далеко латерально. Плоские вдавления на поверхности долек 4 и 5 указывают на границу между червем и полушариями. Эти вдавления наиболее отчетливы в дольке 6 (простой дольке — lobulus simplex, соответствующей скату червя — declive vermis), расположенной каудальнее первой щели. Глубокая парамедианная борозда находится латеральнее дольки 7, но отсутствует в дольке 8 (пирамиде червя — pyramis vermis, соответствущей парамедианной дольке — lobulus paramedianus — и дорсальному околоклочку — paraflocculus dors.) Кора пирамиды продолжается, не прерываясь, в полушарие в виде купола пирамиды — cupula pyramidis. Ни одна из междольковых щелей в каудальной доле, расположенной между первой и препирамидной щелями, не является сплошной между червем и полушарием. У места соединения долек 6 и 7 с полушарием кора мозжечка прерывается и белое вещество выходит на поверхность. Три щели сходятся в этой точке: червячная часть верхней каудальной щели — fiss. caud. sup., расположенная между дольками 6 и 7; полушарная часть этой же щели, отделяющая простую дольку от ножки 1 (ростральной ножки — crus rostrale, соответствующей листку червя — folium vermis) петлеобразной дольки — lobulus ansiformis; межножковая щель петлеобразной дольки. Анзопарамедианная (петлеоколосрединная) борозда, расположенная между каудальным листком ножки 2 {ростральной ножки — crus rostrale, соответствующей бугру червя — tuber vermis) петлеобразной дольки и ростральным листком парамедианной дольки 7, заканчивается в парамедианной борозде латеральнее дольки 7.
Кора дольки 9 (язычка червя — uvula vermis, соответствующего вентральному околоклочку — paraflocculus ventr.) и дольки 10 (узелка — nodulus, соответствующего клочку — flocculus), а также вторая щель — fiss. secunda — и латеральная каудальная щель fiss. caud. lat. — оканчиваются в глубокой парамедианной борозде, которая отделяет эти дольки от купола пирамиды. Латерально купол продолжается в околоклочок.
Срезанный край крыши каудальной части IV желудочка — лента IV желудочка — tenia (taenia) ventriculi quarti — прикрепляется к краю узелка, куполу пирамиды и клочку. Каудальный мозговой парус здесь не заметен, хотя области, свободные от коры, огра ничивающие ленту в парамедианной борозде и вентральную сторону околоклочка, рассматриваются как таковые. Ростральный мозговой парус является продолжением мозжечковых спаек в центральном белом веществе мозжечка. При рассечении мозжечка по стороне мозгового ствола видно, что ростральный мозговой парус продолжает каудальную мозжечковую ножку.
Внутри мозжечка видно его центральное белое вещество с миелиновыми волокнами — мозговое тело — corp. inedullare; на продольном разрезе имеет характерную древовидно ветвящуюся структуру — древо жизни — arbor vitae. Пальцеобразные белые пластинки, проникающие в лепестки мозжечка, называются белыми полосками — laminae albae. Наружную часть червя и полушарий мозжечка составляет серое вещество, образующее кору мозжечка — cortex cerebelli.
Ядра мозжечка — nucll. cerebelli — обычно подразделяются на две группы взаимосвязанных центральных ядер. Каудальная группа состоит из медиального ядра мозжечка (ядро шатра) — nucl. med. cerebelli (nucl. fastigii) и латерального (заднего) вставочного ядра мозжечка — nucl. interpositus lat. (post.) cerebelli, которое соответствует пробковидному ядру — nucl. emboliformis. Ростральная группа включает медиальное (переднее) вставочное ядро мозжечка — nucl. interpositus med. (ant.) cerebelli, соответствующее шаровидному ядру — nucl. globosus, и латеральное ядро мозжечка (зубчатое ядро) — nucl. lat. cerebelli (nucl. dentatus).
IV желудочек —ventriculus quartus — представляет собой полость, принадлежащую всему ромбовидному мозгу, содержащую небольшое количество спинномозговой жидкости и располагающуюся на дорсальной поверхности между продолговатым мозгом и мозжечком. Сводом IV желудочка является червь мозжечка и рос тральный (верхний) мозговой парус — velum inedullare rostralis (sup.) — пластинка между правой и левой ростральными мозжечковыми ножками и язычком мозжечка. Парус соединяется с пластинкой крыши среднего мозга посредством уздечки рострального мозгового паруса — frenulum veli medullaris rostralis. Боковые стенки IV желудочка представлены ножками мозжечка, дно — продолговатым мозгом. Каудально IV желудочек переходит в спинномозговой канал, рострально суживается и переходит в водопровод среднего мозга. Пространство между мозжечком и пластинкой каудального мозгового паруса — lain, velum inedullare caud. — заполнено сосудистым сплетением IV желудочка — pi. choroideus (chorioideus) ventriculi quarti. Вместе с сосудистым сплетением эпителиальная пластинка составляют сосудистую основу IV желудочка — tela oroidea ventriculi quarti, представляющую собой слой мягкой оболочки головного мозга и эпендимы, расположенную между каудальным мозговым парусом и лентой IV желудочка. Дном IV желудочка является ромбовидная ямка — fossa rhomboidea, видимая после удаления мозжечка и крыши IV желудочка. Ямка делится продольной срединной бороздой — sul. medianus; по обе стороны от борозды расположено парное срединное возвышение — eminentia mediana, в область которого проецируются ядра отводящего и подъязычного нервов. От дорсального улиткового ядра тянутся мякотные нервные волокна — мозговые полоски IV желудочка (слуховые полоски) — striae medullaris ventriculi quarti (striae acusticae), которые проходят над каудальной мозжечковой ножкой и идут медиально к срединной борозде дна IV желудочка. Каудомедиальный конец возвышения занимает треугольник подъязычного нерва — trigonum n. hypoglossi, в котором лежит двигательное ядро подъязычного нерва. Латеральнее треугольника подъязычного нерва находится треугольник блуждающего нерва (серое крыло) — trigonum n. vagi (ala cinerea — BNA, JNA), образованный выступающими ядрами блуждающего и подъязычного нервов. На латеральной стороне ромбовидной ямки ростральнее латерального кармана IV желудочка — recessus lat. ventriculus quarti — расположено возвышение медиального вестибулярного ядра — eminentia nucl. vestibularis med., содержащее ядра слухового нерва. Задний суженный конец ромбовидной ямки носит название самого заднего поля (писчего пера) — area postrema (calamus scriptorius — BNA, JNA); является одним из циркумвентрикулярных органов. В каудальной стенке IV желудочка находится непарное поперечное образование — задвижка — obex, которой заканчивается срединная борозда продолговатого мозга. IV желудочек сообщается с подпаутинным пространством при помощи парной латеральной апертуры IV желудочка (отверстия Люшке — Luschka) — apertura lat. ventriculi quarti, расположенной в сосудистой основе IV желудочка в области его латеральных карманов и непарной срединной апертуры IV желудочка (отверстия Мажанди) — apertura lat. ventriculi quarti, расположенной ростральнее задвижки.

Рис. 6 Топография желудочков головного мозга в сагиттальной плоскости: общая схема (А), боковой желудочек (Б), III желудочек (В), желудочко вая система в каудальной части мозгового ствола (Г)
1 — cornu frontale, 2 — pars centralis, 3 — cornu temporale, 4 — pars dors., 5 — recessus pinealis, 6 — nucl. caudatus-putamen, 7 — organum subcommissurale, 8 — соединительная часть, 9 — infundibulum, 10 — pars ventr., 11 — adhesio interthalamica, 12 — colliculus caud., 13 — ventriculus mesencephali, 14 — nucl. fastigii, 15 - can. centralis, 16 — ventriculus quartus, 17 — aqueductus mesencephali.
Спинномозговая жидкость — liquor cerebrospinalis, называется также цереброспинальной жидкостью, ликвором, — является жидкой средой, циркулирующей в полостях желудочков, субарахноидальном пространстве и спинномозговом канале. Образуется в сосудистых сплетениях желудочков мозга, заполняет их полости, а также субдуральное и подпаутинное пространства спинного и головного мозга, сообщающиеся с полостями желудочков посредством отверстий — латеральных и срединной апертур. Отток жидкости за пределы нервной системы происходит через венозную систему твердой мозговой оболочки.
Средний мозг
Средний мозг — mesencephalon — является отделом головного мозга, расположенным между мостом и промежуточным мозгом; представлен ножками мозга, крышей среднего мозга и включает водопровод среднего мозга.
Ножка мозга — pedunculus cerebri — парное образование, расположена вдоль вентролатеральной поверхности среднего мозга; в виде двух сильно развитых валиков идут от полушарий большого мозга к продолговатому мозгу. Ножки разделены вентральной срединной бороздой, которая каудальнее сосцевидных тел образует треугольное пространство — межножковую ямку — fossa interpeduncularis. Дном ямки является каудальное продырявленное вещество — substantia perforata caud., через которое в мозг проходят кровеносные сосуды. Из медиальной борозды ножки мозга — sul. med. crurus cerebri — начинается глазодвигательный нерв. Ножка мозга состоит из вентральной части — основания ножки мозга — basis pedunculi cerebralis, часто называемой ножкой большого мозга — crus cerebri (JNA), и дорсальной части — покрышки среднего мозга — tegmentum mesencephali, разделенных черным веществом.
Крыша среднего мозга — tectum mesencephali, часто именуемая четверохолмием — corpora quadrigemina, — расположена дорсальнее покрышки среднего мозга, представляет собой образование, включающее пластинку крыши, холмики и ручки холмиков. Пластинка крыши — lam, tecti, иногда называемая пластинкой четверохолмия — lam, quadrigemina (BNA, JNA), — состоит из двух пар полушарных возвышений — ростральных и каудальных холмиков. Ростральные холмики — colliculi rostrales, иногда называемые зрительной крышей, или верхними холмиками, — являются подкорковыми центрами зрения; они небольшие, краниально к ним подходит гипофиз, каудально они отделены бороздой от каудальных холмиков — colliculi caud., имеющих также название нижних, или слуховых, холмиков. Обе пары холмиков делятся срединной бороздой на правые и левые. Каждый ростральный холмик посредством ручки рострального холмика — brachium colliculi rostralis — связан с латеральным коленчатым телом. Парная ручка каудального холмика — brachium colliculi caud. — соединяет каудальный холмик с медиальным коленчатым телом. Сзади каудальных холмиков выходят блоковые нервы. Дорсально в каудальный холмик загибается желудочек среднего мозга — ventriculus mesencephali , часто называемый углублением каудального холмика. Желудочек и его эпендимная поверхность окружены околожелудочковым серым веществом, не отличающимся по строению от серого вещества среднего мозга. Общая поверхность желудочка относительно небольшая — 1,4 мм2, что составляет около 2% от общей поверхности желудочковой системы мозга.
Водопровод среднего мозга (водопровод мозга) — aqueductus mesencephali (aquaeductus cerebri, BNА), традиционно называемый сильвиевым водопроводом — aquaeductus Sylvii, — представляет собой узкий канал (расширяющийся каудально), соединяющий III желудочек промежуточного мозга с IV желудочком среднего мозга. По водопроводу происходит отток цереброспинальной жидкости из мозга. В центральной части водопровода, в его базальной стенке выявлены мелкие углубления, увеличивающие эпендимную поверхность; в каудальной части обнаружено одно большое углубление.
В среднем мозгу расположены многочисленные ядра и проводящие пути. Среди них: ретикулярные ядра среднего мозга — nucll. reticulares mesencephali, включающие клинообразное ядро — nuclcuneiformis, ядро плаща — nucl. sagulum и глубокое среднемозговое ядро, центральное серое вещество — substantia grisea centralis, дорсальное и латеродорсальное ядра покрышки — nucll. dors, et laterodors. tegmenti, межножковое ядро — nucl. interpeduncularis и промежуточное ядро — nucl. interstitialis. Кроме того, здесь находятся некоторые ядра черепных нервов: ядро среднемозгового пути тройничного нерва — nucl. tr. mesencephalici n. trigemini, двигательное ядро глазодвигательного нерва — nucl. motorius n. oculomotorii и двигательное ядро блокового нерва — nucl. motorius n. trochlearis. Наконец, в покрышке среднего мозга расположены красное ядро — nucl. ruber и черное вещество — substantia nigra.
Ядра каудального холмика — nucll. colliculi caud. — представляют собой крупные парные образования, занимающие значительную область дорсальной части среднего мозга. Ядра располагаются по обе стороны спайки каудальных холмиков — commissure collicu lorum caudalium — и включают центральное, околоцентральное и наружное ядра (Ryugo, 1976). Центральное ядро — nucl. centralis — у крыс организовано, как и у кошек, в виде пластинок в трех плоскостях. Ядро подразделяется на дорсомедиальную — pars dorsomed. — и вентролатеральную — pars ventrolat. — части. Околоцентральное ядро — nucl. pericentralis — расположено дорсальнее дорсомедиальной части центрального ядра. Наружное ядро — nucl. ext. — представляет собой полоску преимущественно крупных клеток, вставленную между околоцентральным ядром и латеральным краем центрального ядра каудального холмика.
Ростральный (верхний, передний) холмик — colliculus rostralis — или зрительная крыша, представляет собой большое слоистое образование дорсальной и дорсолатеральной частей крыши среднего мозга. Холмик включает следующие слои: поясной слой рострального холмика — str. zonale colliculi rostralis — очень тонкий наружный слой белого вещества; поверхностный серый слой — str. griseum superficiale colliculi rostralis; зрительный слой — str. opticum; промежуточный серый слой — str. griseum intermedium; глубокий серый слой — str. griseum profundum; промежуточный и глубокий мозговые слои — str. medullaria intermedia et profunda.
Предкрыша — pretectum, носит также название предкрышечное (претектальное) поле — area pretectalis, — происходит из эпиталамуса и расположена у наиболее рострального полюса среднего мозга, ограничивая таламус. Хотя область часто рассматривается как единое образование, в настоящее время выявлены несколько предкрышечных ядер — nucll. pretectales, а именно ядро зрительного тракта — nucl. tr. optici, оливное пред крышечное ядро — nucl. pretectalis olivaris, а также ростральное и каудальное предкрышечные ядра — nucll. pretectales rostralis et caud.
Промежуточный мозг
Промежуточный мозг (межуточный) мозг — diencephalon — наряду с конечным мозгом составляет передний мозг; является отделом последнего, расположен под мозолистым телом и сводом, связывает полушария с мозговым стволом. Промежуточный мозг включает гипоталамус, субталамус, таламус, метата ламус и эпиталамус; здесь же рассматривается III желудочек.
III желудочек — ventriculus tertius — непарный желудочек головного мозга, является вторым (после боковых Желудочков) по размеру (27%) среди желудочков мозга. Площадь его поверхности составляет более 20 мм2. Представляет собой полость вокруг межталамического сращения; рострально соединяется с правым и левым боковыми желудочками через межжелудочковое отверстие, каудально — с водопроводом среднего мозга. Разделяется межталамическим сращением на дорсальную и вентральную части. Дорсальная часть расположена над таламусом и составляет около 40% общей поверхности эпендимы III желудочка. В III желудочке различают четыре основных отростка — углубления: зрительное — recessus opticus, нейрогипофизарное, подсосцевидное и шишковидное (или поводковое). Крыша желудочка образована сосудистой основой III желудочка — tela choroidea (chorioidea) ventriculus tertii, которая поддерживает сосудистое сплетение III желудочка — pl. choroideus ventriculi tertii — гирляндоподобное скопление сосудистых ворсинок, идущее от межжелудочкового отверстия до шиш ковидного углубления — recessus pinealis (habenularis). Кроме этого углубления III желудочек заходит в воронку гипофиза, образуя нейрогипофизарное углубление (углубление воронки) — recessus neurohypophysialis (infundibuli). В дорсоростральной части желудочка находится очень маленькая округлая или овальная околожелудочковая непарная структура — субфорникалъный орган (под-сводный орган) — organum subfornicale. Расположен у края межжелудочкового отверстия в месте перехода тела свода в его столбы. Внутри III желудочка на дорсальной, ростральной и вентральной поверхностях каудальной спайки расположен непарный субкомиссуральньш орган — organum subcommissurale, являющийся производным эпендимы III желудочка. III желудочек соединяется с боковым желудочком посредством парного межжелудочкового, или монроева, отверстия — for. interventriculare s. Monroi, расположенного между столбом свода и ростральным концом таламуса. В каудальную стенку III желудочка на границе между средним и промежуточным мозгом входит изогнутая пластинка белого вещества — каудальная спайка (эпиталамическая, или задняя) — commissure caud. (epithalamica s. post.). Этот Х-образный мостик пересекающихся волокон связывает ростральные холмики с контрлатеральными эпиталамическими структурами. Вдоль III желудочка у места соединения гипоталамуса и таламуса проходит гипоталамическая борозда — sul. hypothalamicus.
Гипоталамус — hypothalamus — имеет также названия: гипоталамическая область, подбугорная область, подбугорье, является наиболее вентральной частью промежуточного мозга, расположен вентральнее таламуса и образует дно и стенки III желудочка. Серая конечная пластинка — lam. terminalis grisea — составляет срединную ростральную границу гипоталамуса и ростральную стенку III желудочка. Каудальнее зрительного перекреста — chiasma opticum — дно III желудочка выпячивается вентрально и образует воронку, или воронкообразную часть нейрогипофиза, известную как срединное возвышение. Аденогипофиз, возникающий как дорсальное выпячивание эмбриональной крыши глотки, окружает все части нейрогипофиза и также образует переднюю долю — lobus ant.
Серый бугор — tuber cinereum — является отделом гипоталамуса, его средней частью, расположенной между зрительным перекрестом и сосцевидным телом. В центре серый бугор принимает форму воронки, содержащей в середине нейрогипофизарное углуб ление. Узкая часть воронки анатомически связана с гипофизом. В сером бугре находятся ядра серого вещества.
Сосцевидное тело — corp. mamillare — парное округлое образование в каудальной трети гипоталамуса; расположено ростральнее межножковой ямки и каудальнее серого бугра между двумя нож ками мозга. Представляет собой сложную структуру — сосцевид ный комплекс, в котором под слоем белого вещества содержатся многочисленные ядра. Подсосцевидное углубление — recessus iiifrainamillaris — III желудочка лежит вентральнее комплекса. К латеральным стенкам углубления примыкают каудальные продолжения нескольких ядер серого бугра.
Гипоталамус может быть подразделен на медиальную часть, богатую клетками, илатеральную часть, с меньшим количеством клеток, через которую проходят пучки волокон от конечного мозга к среднему. Большинство хорошо различимых клеточных групп называются ядрами; они окружены более диффузно организованными гетерогенными скоплениями клеток, именуемыми полями (областями). Обычно между смежными полями нет четких границ, например, между медиальным предоптическим и ростральным гипоталамическим полями или между областью серого бугра и латеральным гипоталамическим полем. Гипоталамус подразделяется в рострокаудальном направлении на предоптическую, ростральную, промежуточную (бугорную) и каудальную, или сосцевидную, области, причем дорсальная область находится выше ростральной и промежуточной областей. Соответственно, внутри медиальной части гипоталамуса выделяют медиальную предоптическую, ростральную, промежуточную, каудальную и дорсальную гипоталамическую области. В латеральной части латеральная предоптическая область является каудальным продолжением медиального пучка конечного мозга, связывающего структуры конечного мозга с мозговым стволом.
Медиальная часть гипоталамуса включает ростральную гипоталамическую область с медиальной преоптической областью, промежуточную (серобугорную) гипоталамическую область, каудальную гипоталамическую область и дорсальную гипотала мическую область.
Ростральная (передняя) гипоталамическая область (поле) с медиальной предоптической областью — reg. (area) hypothalamica rostralis (ant.) cum reg. preopticus (prae-) med. — рассматриваются как сплошное образование. Медиальная предоптическая область ограничена рострально ядром диагональной полоски и nucl. accumbens. Она сливается каудально (без хорошо заметной границы) с ростральной гипоталамической областью, которая, в свою очередь, является продолжением (на уровне каудального края зрительного перекреста) промежуточной гипоталамической области. Область включает значительное число ядер и полей. Среди них: срединное и медиальное предоптические ядра — nucll. preoptici medianus et med., ростральное гипоталамическое ядро — nucl. hypothalamic us rostralis супрахиазматическое ядро — nucl. suprachiasmaticus, паравенщрикулярное ядро — nucl. paraventricularis, латеральное и каудальное ростральные ядра nucll. rostrales lat. et caud., дугообразное ядро — nucl. arcuatus, а также медиальное предоптическое поле — area preoptica med. и ростральное гипоталамическое поле — area hypothalamica rostralis.
Промежуточная (серобугорная) гипоталамическая область — reg. hypothalamica intermedia (tuberalis) — является средней частью гипоталамуса, к которой вентрально прикрепляется гипофиз. Как указывалось выше, дно эмбрионального III желудочка выпячивается вентрально и становится нейрогипофизом. В промежуточной области различают несколько полей и ядер. Среди них: ретрохиазматическое поле — area retrochiasmatica и серобу горное поле — area tuberale, дорсальное и гигантоклеточное ядра серого бугра — nucll. tuberis dors, et inagnocellularis, а также гипоталамические ядра: вентромедиальное — nucl. hypothalamicus ventromed., dopcoмедиальное — nucl. hypothalamicus dorsomed. и перифорникальное — nucl. hypothalamicus perifornicalis.
Каудальная гипоталамическая область — reg. hypothalamica caud., называемая также задней, или сосцевидной, областью, входит в состав каудальной трети гипоталамуса, наряду с сосцевидным комплексом и двумя гигантоклеточными группами. Сосцевидное углубление III желудочка расположено вентральнее сосцевидного комплекса. К латеральным стенкам углубления прилежат каудальные продолжения нескольких ядер серого бугра: дугообразного, дорсомедиального ядер и гигантоклеточного ядра серого бугра, и более каудально — дорсальное и вентральное предсосцевидное ядра и каудальное гигантоклеточное ядро сходятся над улублением. Область включает каудальное гипоталамическое поле — area hypothalamica caud. и множество ядер сосцевидного комплекса: вентральное и дорсальное предсосцевидные ядра — nucll. preinammilares ventr. et dors., медиальное и латеральное сосцевидные ядра — nucll. maminilares med. et lat., подсосцевидно-таламическое — nucl. submammilothalamicus и надсосцевидное ядро — nuclsuprainammilaris. Кроме того, здесь расположены каудальное гигантоклеточное ядро — nucl. inagnocellularis caud. и близнецовые ядра — nucll. gemini.
Дорсальная гипоталамическая область — reg. hypothalamica dors. — расположена над ростральной областью и серым бугром и тянется рострально от каудовентральной границы ростральной спаики до каудальной гипоталамической области и среднего мозга Kaудально. Латерально область ограничивается сводом и латеральной гипоталамической областью, дорсальнее ее лежат таламус и сосце видно-таламический пучок. Область сосуществует с дорсальным гипоталамическим полем — area hypothalainica dors., в котором расположено дорсальное гипоталамическое ядро — nucl. hypothalamicus dors. Большую часть области занимает дорсальная часть паравентрикулярного ядра — nucl. paraventricularis (pars dors.).
Латеральная часть гипоталамуса расположена латераль нее его медиальной части. Латеральная пред оптическая область (рострально) и латеральная гипоталамическая область составляют одно скопление сгруппированных клеток, которое сосуществует с латеральной частью. Топографически латеральная часть подразделяется рострокаудально на предоптическо-ростралъную, бугорную и каудальную, или сосцевидную, области, соответствующие их прилежащим медиальным областям. В этих областях располагается небольшое число хорошо очерченных ядер. Латеральная предоптическая и латеральная гипоталамическая области обеспечивают расположение большого пучка проходящих волокон, известного как медиальный пучок конечного мозга — fasc. telencephalicus med., соединяющий структуры конечного мозга и мозгового ствола.
Ростральная гипоталамическая область с предоптической областью включают латеральное предоптическое поле — area preoptica lat. и супраоптическое ядро — nucl. supraopticus. Само ядро и его ретрохиазматическая часть — pars retrochiasmatica имеют мощные связи к нейрогипофизу. Нейроны ядра продуцируют окситоцин и аргинин-вазопрессин.
Промежуточная гипоталамическая (бугорная) область включает латеральное гипоталамическое поле — area hypothalainica lat., латеральное и медиальное ядра серого бугра — nucll. tuberis lat. et med.
Каудальная гипоталамическая область включает латеральное гипоталамическое поле, идущее каудально, латеральнее сосцевидных ядер и рострального полюса медиального сосцевидного ядра; Далее поле замещается вентральным полем покрышки среднего мозга.
Поперечные пучки нервных волокон, расположенные над зрительным перекрестом и соединяющие структуры левой и правой половины мозга, называются супраоптическими спайками — соmmissurae supraopticae. Дорсальная супраоптическая спайка, или спайка Ганзера, — commissure supraoptica dors. — содержит перекрещивающиеся волокна от медиального бледного шара. Вентральная супраоптическая спайка — commissure supraoptica ventr. — включает две спайки — верхнюю спайку, или спайку Мейнерта, — сommissura sup. (— Meynerti — BNA) — и нижнюю спайку, или спайку Гуддена, — commissure inf. (— Guddeni — BNA).
Проекционные нервные пути — tr. nervosi projectiones, соединяют клетки коры полушерий большого мозге с клеткеми нижерасположенных отделов головного мозга и спинного мозга; различают
восходящие и нисходящие пути. Существует два основных пути афферентных волокон к сосцевидным ядрам — свод и сосцевидная ножка.
Свод — fornix — представляет собой совокупность двух изогнутых тяжей белого вещества, расположенных вентральнее мозолистого тела, соединенных в средней части в виде непарного тела свода — corp. fornicis, а рострально и каудально расходящихся образуя соответственно столбы и ножки свода.
Сосцевидная ножка — pedunculus mainillaris, иногда называется ножкой сосцевидного тела — pedunculus corporis mainillaris, является пучком волокон, расположенным латеральнее межножковой ямки; содержит волокна от дорсального и вентрального ядер покрышки к латеральному сосцевидному телу и ростральной части медиального сосцевидного ядра.
Наряду со сводом и сосцевидной ножкой в гипоталамусе четко прослеживаются проекционные пути лимбической системы — со сцевидно-таламический — tr. mamillothalamicus и сосцевидно-покрышечный — tr. mainiilotegmentalis пути.
Субталамус — subthalamus — небольшая структура промежуточного мозга, расположена вентральнее таламуса и ростральнее черного вещества. Состоит из субталамического тела — corp. subthalamicus, включающего субталамическое ядро — nucl. subthalamicus и прилегающие латеральные структуры, и неопределенной зоны — zona incerta. Ростральнее субталамического ядра расположено внутриножковое ядро — nucl. entopeduncularis.
Таламус — thalamus, имеет также название зрительных бугров (чертогов), является частью промежуточного мозга, представляющую собой парное скопление серого вещества. Таламус и связанные с ним структуры (метаталамус и эпиталамус) объединяются в понятие таламический мозг (зрительный мозг) — thalamencephalon. Таламус вместе с эпиталамусом (комплекс поводка и шишковидное тело) составляют дорсальную часть промежуточного мозга, а его вентральная часть образована субталамусом. Таламус крысы представляет собой тип, характерный для млекопитающих. Он включает парную ядерную массу, и у крысы, в отличие от хищных и приматов, отсутствует частичное разделение правого и левого таламуса III желудочком. Вместо этого два таламуса простираются через срединную линию и так образуют большое межталамическое сращение (промежуточную массу) — adhesio interthalamica (inassa intermedia) — между дорсальными областями промежуточного мозга. Ростральная, латеральная и прилежащая вентральная поверхности таламуса ограничены и отделены от внутренней капсулы тонким слоем белого вещества, составляющим наружную мозговую пластинку таламуса — lain, inedullaris thalaini ext., и ретикулярным ядром таламуса. На каудальных уровнях поверхность таламуса частично отделена от подлежащих субталамуса и гипоталамуса при помощи наружной мозговой пластинки, медиальной петли и других систем волокон.
Таламус изнутри подразделяется при помощи пластинки миелинизированных волокон — внутренней мозговой пластинки та ламуса — lam. inedullaris thalami int. — на ростральную, медиальную и латеральную части, которые содержат разнообразные группы ядер. Ростральная группа ядер образует ростральную часть, а медиальная часть занята дорсомедиальным ядром таламуса — nucl. dorsomed. thalami. Латеральная часть расположена между наружной и внутренней мозговыми пластинками таламуса и включает вентральную, каудальную и латеральную ядерные группы. Внутрипластинчатая ядерная группа является вставочной во внутренней мозговой пластинке, и по срединной линии эта группа ядер, не прерываясь, переходит в срединную ядерную группу. Наконец, комплекс поводка эпиталамуса лежит дорсальнее медиальной части таламуса, примыкая к латеральной стенке III желудочка.
Ростральные (передние) ядра таламуса — nucll. rostrales (ant.) thalami — включают три ядра: дорсальное ростральное — nucl. rostralis dors., вентральное ростральное — nucl. rostralis ventr. и медиальное ростральное — nucl. rostralis med.
Вентральные ядра таламуса —nucll. ventr. thalami — также называются вентральным ядерным комплексом, или вентральными ярусными ядрами, являются наибольшим комплексом ядер в дорсальном таламусе. Эти ядра составляют 25% объема дорсального таламуса и занимают почти полные вентролатеральные 2/3 большой области, расположенной между наружной и внутренней мозговыми пластинками таламуса. Вентральные ядра подразделяются на медиальное, латеральное и каудальное ядра. Последнее, в свою очередь, включает медиальную, латеральную и мелкоклеточную части.
Медиальное вентральное (вентромедиальное) ядро — nucl. ventr. med. (ventromed.) — составляет 15% от вентрального ядерного комплекса. Оно является продольно вытянутым ядром,занимающим вентромедиальную часть комплекса. На фронтальном срезе ядро треугольной формы и имеет медиальную, вентральную и дорсолатеральную поверхности.
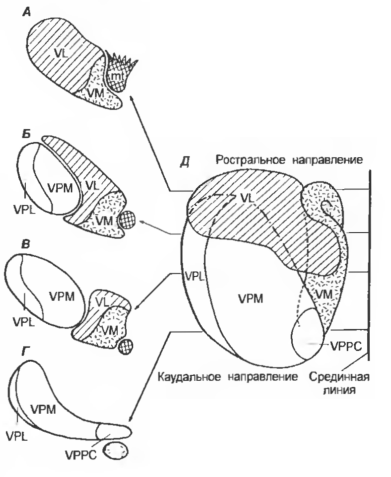
Рис. 7 Взаимоотношении ядер вентрального и вентрокаудального комплексов таламуса на фронтальных рострокаудальных срезах (А-Г) и на схеме с дорсальной стороны (Д)
VPM — nucl. ventr caud. (pars med.), VPL — nucl. ventr. caud. (pars lat.), VL — nucl. ventr. lat. (ventrolat.), VM — nucl. ventr. med., VPPC — nucl. ventr. caud. parvocellularis, mt — tr. niamillotha lamicus.
Латеральное вентральное (вентролатеральное) ядро — nucl. ventr. lat. (ventrolat.) — составляет четверть вентрального комплекса и расположено в ростральной его половине. Вначале ядро появляется на срединных срезах таламуса в виде дорсолатерального продолжения, схожего по клеточному составу с медиальным вентральным ядром. Оно расположено в промежутке, ограниченном медиальным ядром медиально, латерально — медиальной и латеральной частями каудального ядра, дорсально — паравентрикулярным ядром.
Каудальные (задние) вентральные ядра — nucll. ventr. caud. (ventropost.) — являются наибольшей (образуют 60% всего вентрального комплекса) и наиболее четко очерченной составляющей вентральных ядер. В дорсальной проекции они выглядят как крупная овальная ядерная масса, занимающая каудолатеральные 2/3 вентрального ядерного комплекса. Каудальные вентральные ядра включают медиальную и латеральную части и мелкоклеточное ядро.
Каудальное вентральное ядро, медиальная часть (заднемедиальное ядро) — nucl. ventr. caud., pars med. (nucl. ventroposteromed.) — является наибольшим (составляет 57% всего комплекса) и наиболее заметным среди каудальных ядер. Эта часть образует дорсомедиальные 2/3 комплекса, рострально она ограничена латеральной частью каудального вентрального ядра и латеральным вентральным ядром. Она увеличивается и тянется каудально, образуя каудальный полюс вентрального ядерного комплекса.
Каудальное вентральное ядро, латеральная часть (заднелатеральное ядро) — nucl. ventr. caud., pars lat. (nucl. ventroposterolat.) — составляет 40% каудального вентрального комплекса; она занимает наиболее латеральную и ростральную области комплекса и ограничена латерально наружной мозговой пластинкой таламуса, медиально — латеральным вентральным ядром и медиальной частью каудального вентрального ядра.
Мелкоклеточное каудальное вентральное ядро — nucl. ventr. caud. parvocellularis, иногда рассматривается как подразделение медиальной части каудального вентрального ядра, ошибочно называется «базальным вентральным комплексом», или описывается как компонент медиального вентрального комплекса под термином «базальное вентромедиальное ядро». Представляет собой мелкое прямоугольной формы ядро, расположенное в каудальной и медиальной частях каудального вентрального комплекса.
Латеральные ядра — nucll. lat. — у крысы включают каудальное (заднее) латеральное ядро — nucl. lat. caud. (lateropost.), рас
положенное медиальнее и каудальнее дорсальной части латерального коленчатого тела и рассматриваемое как гомолог подушки — pulvinar — таламуса у приматов, и дорсальное латеральное ядро — nucl. lat. dors, (laterodors.).
Каудальные (задние) ядра — nucll. caud. (post.) — относительно большая, трудно различимая область. Ядра расположены дорсомедиальнее медиальной части каудального вентрального ядра, нейтральнее каудального латерального ядра и латеральнее внутренней мозговой пластинки. Ранее область каудальных ядер у крыс состояла из очень маленького поля на уровне перехода среднего мозга в промежуточный и обозначалась как медиальная гигантоклеточная часть медиального коленчатого тела.
Внутрипластинчатые ядра таламуса — nucll. intralaminares thalaini — расположены во внутренней мозговой пластинке таламуса. Они включают ростральную группу из трех ядер: медиального и латерального центральных ядер — nucll. centrales med. et lat. и парацентрального ядра — nucl. paracentralis, и каудального парафасцикулярного ядра — nucl. parafascicularis. Ростральная группа ядер образует сплошной неплотный ядерный комплекс, огораживающий вентральную и латеральную стороны дорсомедиального ядра. Каудально они переходят в большое парафасцикулярное ядро, лежащее в каудальной трети таламуса.
Медианные ядра таламуса — nucll. mediani thalami, иногда называются ядрами срединной линии, образуют две группы: дорсальную — состоит из парных паравентрикулярного— nucl. paraventricularis — и окололенточного — nucl. parataenialis — ядер и непарных межмедиалъноростралъного — nucl. intermediorostralis — и междорсальномедиального — nucl. interdorsomed. ядер, перекрывающих срединную линию, и вентральную — включает главным образом непарные ромбовидное — nucl. rhomboideus — и соединяющее — nucl. reuniens ядра. Дорсальная и вентральная группы отделены друг от друга по всей рострокаудальной протяженности телом и проксимальными частями крыльев медиального центрального ядра. Медианные ядра рассматриваются как вставочные ядра таламической (дорсальной) околожелудочковой системы волокон.
Метаталамус — metathalamus, иногда называется забугорной областью, или забугорьем, является частью таламического мозга, включающей латеральное и медиальное коленчатые тела.
Латеральное коленчатое тело — corp. geniculatum lat. — является парным образованием промежуточного мозга, расположенным в латеральной оконечности таламуса, связано с ростральным холмиком. В латеральном коленчатом теле оканчивается латеральный корешок зрительного тракта. Основную часть коленчатого тела занимает латеральное коленчатое ядро — nucl. geniculatus lat., подразделяющееся на дорсальную — pars. dors, и вентральную — pars, ventr. части.
Медиальное коленчатое гело — corp. geniculatum med. — является парной структурой промежуточного мозга, связано с каудальным холмиком. Основную, клеточную часть коленчатого тела составляет медиальное коленчатое ядро — nucl. geniculatus med., очень слабо изученное у крыс. Ядро подразделяется на три части: вентральную — pars ventr., дорсальную — pars dors, и медиальную — pars med.
Эпиталамус — epithalamus, иногда называется надбугорьем, или надталамической областью; является частью таламического мозга, включающей шишковидную железу — corp. pineale, поводки и связанные с ними структуры.
Поводок — habenula — парное образование белого вещества, прикрепляющее шишковидную железу к таламусу. Симметричные поводковые ядра — nucll. habenulares левой и правой сторон соединяются перемычкой, содержащей перекрещенные нервные волокна — спайкой поводков — commissure habenularum. Ростральным продолжением поводка является большой тяж мякотных нервных волокон — поводковая (мозговая) полоска таламуса. Она рассматривается как перекрещивающийся нервный путь для структур, связанных с поводком.
Конечный мозг
Конечный мозг — telencephalon — представляет собой наиболее развитую часть переднего мозга и по величине больше всех отделов центральной нервной системы. Основными составляющими конечного мозга являются большой мозг, обонятельный мозг и полосатое тело; здесь же рассматриваются боковые желудочки.
Большой мозг — cerebrum — включает два полушария, соединенные спайками. Глубокая продольная щель большого мозга — fiss. longitudinalis cerebri — разделяет его на два полушария. В глубине продольной щели левое и правое полушария соединяются между собой широким поперечным слоем белых волокон — мозолистым телом. В каждом полушарии вентральную часть образует обонятельный мозг, дорсальную — кора больших полушарий. Оба полушария отделяются от мозжечка поперечной щелью большого мозга — fiss. transversa cerebri. Вентральная сторона мозга называется основанием мозга — basis cerebri.
Полушарие — hemispherium (hemisphaerium) парное симметричное и крупнейшее образование мозга. В каждом полушарии различают выпуклую (дорсолатеральную) поверхность — facies convexa (dorsolat.), медиальную поверхность — facies med. — и базилярную, или вентральную, поверхность — facies basilaris. Выделяют также дорсальный (лобный) полюс— polus rostralis (frontalis) — и каудальный (затылочный) полюс — polus caud. (occipitalis). Полушария покрыты слоем серого вещества — корой большого мозга, или плащом, — cortex cerebri s. pallium.
Как и у всех млекопитающих, у крысы кора подразделяется на изокортекс и аллокортекс. Изокортекс — isocortex, имеет также названия: новая кора — neocortex, неокортекс, гомогенетическая кора (Brodmann, 1909); является частью коры большого мозга, проходящей в эмбриогенезе стадию шестислойного строения; филогенетически наиболее поздняя часть коры. Аллокортекс — allocortex — имеет также название гетерогенетическая кора — часть коры полушарий большого мозга, которая в онтогенезе не подразделяется на слои; филогенетически более ранняя часть коры, включает древнюю и старую кору. Древняя кора — paleocortex (palaeo) — является корой базальной части обонятельного мозга; филогенетически наиболее ранняя часть коры большого мозга. Старая кора — archicortex — представляет собой кору перегородочной и лимбической частей обонятельного мозга (гиппокамп, зубчатая извилина); возникает в филогенезе позже древней, но ранее новой коры. Изокортекс и аллокортекс отделены друг от друга переходными корковыми областями (межуточной корой) с преобладанием изокортекса — произокортекс или аллокортекса (периаллокортекс — periallocortex). Последний подразделяется на промежуточную старую кору, или периархикортекс, — mesoarchicortex s. periarchicortex — и промежуточную древнюю кору, или перипалеокортекс, — mesopaleocortex s. peripaleocortex. Распределение участков изо-, алло- и периаллокортекса. Изокортекс покрывает большую часть дорсальной, значительную часть латеральной и небольшие участки медиальной и базальной сторон мозга. Аллокортекс и соответствующие переходные области полностью окружают изокортекс. Разграничение областей произокортекса от изокортекса у крыс очень сложное.
Рис. 8
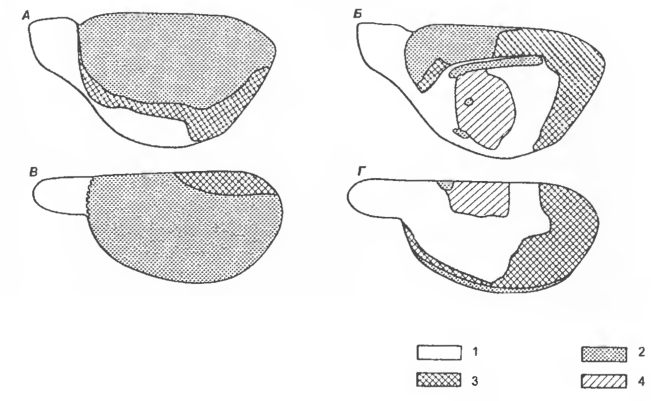
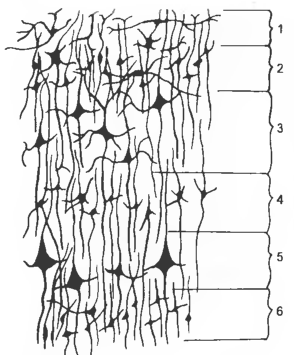
Рис. 9 Поперечный гистологический срез новой коры
1 — str. moleculare (plexiforme), 2 — str. granulare ext., 3 — str. pyramidale ext., 4 — str. granulare int., 5 — str. pyramidale int., 6 — str. multiforme.
Новая кора состоит из шести слоев, содержащих тела нервных клеток и нервные волокна. Выделяют молекулярный, наружный и внутренний зернистые, наружный и внутренний пирамидные и мультиформный слои. Молекулярный слой (пластинка) — str. (lam.) moleculare (plexiforme) — является наружным цитоархитектоническим слоем коры полушарий; содержит ветвящиеся афферентные волокна, тангенциальные нервные волокна — neurofibrae tangentiales, окончания дендритов пирамидных клеток, глиальные элементы и редкие тела нервных клеток. Наружный зернистый слой (пластинка) — str. (lam.) granulare ext. — расположен между молекулярным и наружным пирамидным слоями; образован малыми зерновидными, звездчатыми и пирамидными нейронами и их дендритами. Наружный пирамидный (пирамидальный) слой (пластинка) — str. (lam.) pyramidale ext. — слой коры большого мозга, находящийся между наружным и внутренним зернистыми слоями. Состоит из средних и крупных пирамидальных нейронов, их дендритов и первого сегмента аксонов; наиболее широкий слой неокортекса. Внутренний зернистый слой (пластинка) — str. (lam.) granulare int. — расположен между наружным и внутренним пирамидными слоями. Содержит специфические таламические афферентные волокна, малые зерновидные, звездчатые и пирамидные нейроны. В сенсорных полях коры эти нейроны упакованы, образуя толстый слой. Внутренний пирамидный слой (пластинка) — str. (lam.) pyramidale int. — находится между внутренним зернистым и мультиформным слоями; образован преимущественно средними и крупными пирамидами, в том числе гигантопирамидными нейронами. Мультиформный (полиморфный) слой (пластинка) — str. (lam.) multiforme самый внутренний слой коры большого мозга, образованный разнообразными нейронами, преимущественно веретенообразной формы; иногда присутствуют мелкие и средние пирамидные клетки.
Номенклатура корковых областей. Начиная с 20-х годов XX в. крыса использовалась в многочисленных исследованиях по изучению роли отдельных областей коры в поведенческих реакциях. Однако только в конце 40-х годов Кригом (Krieg) было
проведено первое полное описание корковых областей мозга крысы, основанное на приложении цифровой номенклатуры Бродмана (Brodmann, 1909) для коры большого мозга человека к мозгу крысы. Эта номенклатура была разработана главным образом на топографической схожести областей у приматов и грызунов, но не на основе цитоархитектонических сравнений. Дальнейшим шагом в этом направлении стал стереотаксический атлас коры мозга крысы, разработанный Зиллесом (Zilles, 1985). В нем предусмотрена номенклатура, независимая от разработанных ранее на других видах животных. Буквенные сокращения используются для обозначения больших областей коры (Par — parietalis — теменная, Те — temporalis — височная и т. д.) или для обозначения традиционно используемых терминов (AI — agranularis insularis — агранулярная островковая).
В дополнение к буквенным обозначениям используются и цифровые, указывающие на определенные функциональные подобласти (напр.: Ос1 и Ос2 означают первичное и вторичное зрительные поля). В табл. 1 дано сопоставление корковых полей (Krieg) с областями коры (по Zilles).
Многие из этих областей коры могут быть включены в предполагаемые функциональные объединения. Предлобная (префронтальная) кора включает все образования, получающие связи от дорсомедиального ядра таламуса. Кора разделяется на несколько подобластей на основании их архитектоники и связей. Выделяют ростральную поясную кору (включающую Cg1, Cg2, Cg3), «область плеча» (Fr2), являющуюся предположительно гомологом дугообразной коры (поле 8 по Бродману) у обезьян, глазничное предлобное поле (латеральное, медиальное и вентролатеральное поля) и ростральную периринальную (околоносовую) область агранулярной островковой коры (AI). Инфралимбическое поле также включено в предлобную кору так как показано, что оно получает афферентные связи от дорсомедиального ядра таламуса. Двигательная (моторная) кора объединяет главным образом первичную моторную кору (.Fr1, Fr3), а также области представительства грудной (FL) и тазовой (HL) конечностей. Соматосенсорная кора охватывает поля 1 (Par1) и 2 (Раг2) теменной коры, представляющие собой первичную и вторичную соматосенсорные «ratunculi», описанные Woolsey (1958). Висцеральная кора (Vi) является самостоятельной структурой, о связях которой очень мало известно, но можно считать доказанным ее сходство с Par1 и Раг2. Зрительная кора традиционно у крысы включает поля 17, 18, и 18а (по Krieg), которые были подразделены Zilles на несколько областей: Oc2L, Oc2ML, Ос2ММ — латеральная, медиолатеральная и медиомедиолатеральная части поля 2, соответственно. В зрительную кору также включается поле 2 височной коры (Те2). Слуховая кора включает поле 1 височной коры (Те1) и, возможно, поле 3 (Те3), которые оба связаны с медиальным коленчатым ядром. Вероятно, что поле 2 височной коры (Те2) также участвует в слуховой функции, так как получает связи от медиального коленчатого ядра в дополнение к связи от каудального латерального ядра таламуса. Каудальная (задняя) ассоциативная кора приблизительно соответствует полю 7 (по Krieg) и идентифицируется с задней теменной корой (Kolb, Wolkey, 1987). Согласно атласу Zilles нет различий между возможной задней теменной корой и областями Ос2М и Oc2L. Ассоциативная островковая кора включает периринальное поле (PRh) и поля 2 (Те2) и 3 (Те3) височной коры.

Рис. 10 Карта новой коры с прилежащими областями аллокортекса, составленная по материалам цито- и миелоархитектонических исследований: латеральная (А), дорсальная (Б) и медиальная (В) поверхности Fr1-Fr3 — агеае frontales, Par1-Par2, FL, HL — areae parietales, Tel-Te3 — areae temporaries, Oc1B, Oc1M, Оc2L, Oc2MM, Oc2ML — areae occipitales, Cg-Cg3 — cortex prefrontalis med., IL — area infralimbica, MO, LO, VO — cortex orbitalis, AID, AIP, ATV — cortex insularis, PRh — cortex perirhinalis, RSA, RSG — cortex retrosplenialis; аллокортекс и части периаллкортекса обозначены квадратной штриховкой, переходные участки диэнцефалон-теленцефалон косой штриховкой.
Аллокортекс включает древнюю и старую кору. На медиальной и дорсальной поверхностях полушарий границу между аллокортексом и изокортексом образуют ростральная поясная кора (поля Cgl-3 и инфралимбическое поле медиальной лобной коры — IL) и продолжающаяся каудально ретросплениальная кора (агранулярные поля RSA и 29D, гранулярное поле RSG). На латеральной и ростральной поверхности изокортекс ограничен агранулярной островковой корой (или клаустрокортексом), которая подразделяется на дорсальную (AID), вентральную (AIV) и заднюю (А1Р) части. Каудально, где за грушевидной корой следует энторинальная кора, периринальная кора (PRh) образует латеральную границу изокортекса. Строение грушевидной коры, миндалевидного тела, а также образований гиппокампальной области будут представлены при описании структур обонятельного мозга.
Изокортекс является синонимом для обозначения неокортекса, или новой коры. Номенклатура областей новой коры часто основывается на результатах, полученных на приматах, включая человека. Поэтому в работах иногда могут использоваться термины классификации коры по Бродману. Krieg (1946, 1947) применил для крысы цифровую номенклатуру Бродмана для коры мозга человека без достаточного соответствия.

Рис. 11 Области коры с латеральной (А) и дорсальной (Б) поверхностей Список принятых сокращений участков коры представлен в табл. 1.
Другая терминология основана на функциональной схожести корковых областей приматов и грызунов. Поэтому Zilles разработана исчерпывающая, нейтральная и независимая номенклатура неокортекса крысы. Новая кора разделяется топографически на лобную (Fr), теменную (Par), височную (Те) и затылочную (Ос) области. Каждая область подразделяется на несколько полей, обозначенных цифрами. Некоторые поля, в свою очередь.

Рис. 12 Области коры с медиальной (А) и базальной (Б) поверхностей Список принятых сокращений участков коры представлен в табл. 1.

Разделяются дальше, например поле 2 затылочной коры (Ос2) на латеральную (Oc2L), медиолатеральную (Oc2ML) и медиомедиальную (Ос2ММ) части.
Обонятельный мозг — rhinencephalon — является частью конечного мозга; состоит из базального конечного мозга, гиппокампа и ассоциированных образований. Подразделяется на три части: базальную, перегородочную и лимбическую.
Базальная часть обонятельного мозга — pars basal is rhinencephali — включает основную и добавочную обонятельные луковицы, ростральное обонятельное ядро, грушевидную кору и другие образования.
Обонятельная луковица — bulbus olfactorius является наиболее ростральной частью обонятельного мозга. Лежит сразу же напротив продырявленной решетчатой пластинки (решетчатой кости), которая разделяет носовую и черепную полости. От луковиц в каудальном направлении по вентральной поверхности мозга идут тонкие тяжи волокон латерального обонятельного тракта — tr. olfactorius lat., который проецируется к различным обонятельным областям переднего мозга. Медиально по сторонам обонятельных трактов расположен парный обонятельный бугорок — tub. olfactorium, имеющий три слоя: полиформный — str. polyforme, пирамидный — str. pyramidale — и сетчатый — str. plexiforine.
В обонятельной луковице хорошо различимы шесть слоев: слой обонятельного нерва — str. n. olfactorii, клубочковый — str. glomerulare, наружный сетчатый — str. plexiforme ext., слой митральных клеток — str. mitrale, внутренний сетчатый — str. plexiforme int. и внутренний зернистый слой — str. granulare int.
Добавочная обонятельная луковица — bulbus olfactorius accessorius — расположена дорсально и несколько медиально, между главной обонятельной луковицей и ростральным обонятельным ядром — nucl. olfactorius rostralis — в виде чечевицеобразного включения в обонятельную луковицу. Добавочная обонятельная луковица у крыс, как и у других грызунов, зайцеобразных и насекомоядных, имеет внутренний сетчатый слой большой толщины. Фактически, этот слой состоит из широкой ленты белого вещества — дорсального латерального обонятельного тракта — tr. olfactorius lat. dors.
Грушевидная (пириформная) кора — cortex piriformis — представляет собой древнюю кору ростральной части грушевидной доли, известную также как первичная обонятельная кора, или предгрушевидная кора. Название грушевидная доля — lobus piriformis — традиционно использовалось как общий термин для рострального обонятельного ядра, первичной обонятельной (или грушевидной) коры и энторинальной области, которые образуют структуру более или менее грушевидной формы, по крайней мере у макросматиков, как крыса. Рострально кора тянется в обонятельный бугорок, где она продолжается в латеральную переходную зону рострального обонятельного ядра. Каудально она постепенно замещается энторинальной областью (периаллокортекс). Грушевидная кора ограничена латерально от лобной и островковой коры носовой бороздой, а медиально от обонятельного бугорка и поверхностного миндалевидного тела — дугообразной и миндалевидно) бороздами, соответственно. Кора подразделяется на ростральную и каудальную части поперечной линией, проведенной через каудальный полюс обонятельного бугорка. Часть грушевидной коры, прилегающая к миндалевидному телу, иногда называется периамигдалярной корой.
Рис. 13 Обонятельные проекционные области мозга с вентральной поверхности
1 — tub. olfactorium, 2 — cortex piriformis, 3 — nucl. amygdaloideus corticalis rostralis, 4 — nucl. amygdaloideus corticalis caudolat., 5 — cortex entorhinalis lat., 6 — cortex entorhinalis med., 7 — nucl. amygdaloideus corticalis caudomed., 8 — nucl. amygdaloideus med., 9 — nucl. proprius tr. olfactorii accessorii, 10 — nucl. tr. olfactorii lat., 11 — tr. olfactorius lat., 12 — bulbus olfactorius.
Перегородочная часть обонятельного мозга —par septalis rhinencephali — находится на медиальной стороне полуша рия и представлена в основном перегородкой конечного мозга.
Перегородка конечного мозга — septum telencephali, носит на звание прозрачной перегородки — septum pellucidum (термин применяется для млекопитающих-микросматиков) — или клеточной (истинной) перегородки — septum cellulare (verum); последние синонимы используются для низших млекопитающих, у которых толстая перегородка содержит много нервных клеток. Перегородка соединяет мозолистое тело со сводом и разделяет боковые желудочки. Перегородка является собранием ядер и пучков волокон. Hа основании изучения топографии, цитоархитектоники и связей области перегородки (септальной области) она подразделена на латеральную, медиальную, каудальную и вентральную области.
Латеральное ядро перегородки — nucl. lat. septi — занимает всю латеральную область и подразделяется на дорсальную — pars dors., промежуточную — pars intermedia и вентральную — pars ventr. части.
Медиальная область перегородки включает перегородочно-гиппокампальное ядро — nucl. septohippocampalis, медиальное ядро перегородки — nucl. med. septi и ядро диагональной полоски (Брока) — nucl. tr. diagonalis.
В каудальной области перегородки расположены Две основные клеточные группы — бахромчатое ядро перегородки — nucl. fimbrialis septi и треугольное ядро перегородки — nucl. triangularis septi.
Две группы клеток составляют вентральную область перегородки — перегородочно-гипоталамическое ядро — nucl. septohypothalamicus и вставочное ядро терминальной полоски — nucl. interstitialis striae terminalis.
Лимбическая часть обонятельного мозга — pars mbica rhinencephali — является краевой (limbus — кайма, ободок) и включает структуры вокруг внутренней границы коркового плаoа — гиппокамп и миндалевидное тело.
Гиппокамп — hippocampus — парное образование, относящееся к старой коре и расположенное на медиальной стенке височного бокового желудочка. Является центральной структурой лимбической системы мозга (включающей, кроме того, миндалевидное тело, перегородку, обонятельную луковицу, свод и сосцевидные тела). Гиппокампальная область крысы является одной из наиболее хорошо изученных структур мозга. Она занимает большую часть вентрокаудальной и вентролатеральных стенок коры большого мозга. В направлении от носовой щели (RF) к медиальному краю коры различаются: энториналъная кора — cortex entorhinalis (Ent), околопод-ставка — parasubiculum (PaS), предподставка — presubiculum (PrS), собственно подставка — subiculum (S), поля CA1-CA3 —hippocampus CA1-САЗ — ножки гиппокампа, зубчатая извилина — gyrus dentatus (DG). Гиппокамп (ножка и зубчатая извилина) тянется рострально ниже мозолистого тела к каудальному уровню ядер перегородки. В гиппокампе различают три части: ретро-, супра- и предкомиссуральную части.
В состав гиппокампа входят ножка гиппокампа (аммонов рог) — pes hippocampi (cornu Ammonis) (Amn) — вентральный конец гиппокампа, лоток гиппокампа — alveus hippocampi» представляющий собой тонкий слой белого вещества, покрывающий внутреннюю поверхность гиппокампа, и бахромка гиппокампа — fimbria hippocampi (fi), являющаяся свободной каймой лотка, идущей вдоль границы гиппокампа параллельно зубчатой извилине; продолжается далее в виде ножки свода.
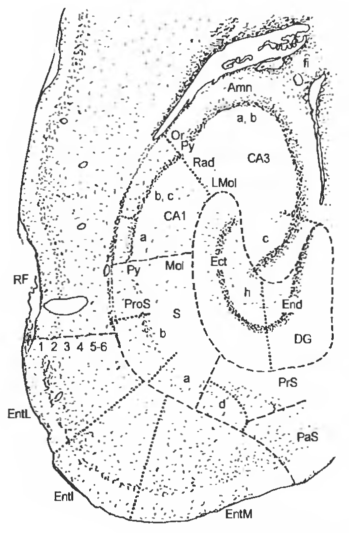
Рис. 14 Микрофотография горизонтального среза области гиппокампа
Принятые сокращения участков коры представлены в тексте, пунктирными линиями ограничены шесть основных структур гиппокампальной области, точечными линиями выделены части этих структур; 1-6 — слои височной коры.
Спайки гиппокампа относятся к межполушарным спайкам, наряду с ростральной спайкой и мозолистым телом. Различают вентральную спайку свода (гиппокампа ) — commissura formcis (hippocampi) ventr. и дорсальную спайку свода (гиппокампа) — commissura fornicis (hippocampi) dors.
Миндалевидное тело — corp. amygdaloideum, часто называется миндалина, или амйгдала, — amygdala — хотя является частью базальных ядер, функционально связано с обонятельным мозгом. Представляет собой структурно организованный комплекс ядер, расположенный под корковым слоем нижневисочной доли мозга. Традиционно миндалевидное тело подразделялось на корково медиальную (обонятельную) часть — pars corticomed. (olfactoria) — и базальнолатеральную часть — pars basolat. Современные достижения гистохимии и определение связей различных структур позволили разделить миндалевидное тело на обонятельную миндалину, медиальную, базально-латеральную и центральную миндалевидную группы.
Обонятельная миндалина включает ядро латерального обонятельного тракта, ростральное миндалевидное поле, ростральное и каудолатеральные корковые миндалевидные ядра, амигдалопириформное переходное поле и ростральный конец медиального миндалевидного ядра.
Медиальная миндалевидная группа объединяет медиальную миндалину, каудомедиальные части подчечевицеподобного безымянного вещества, а также медиальную и промежуточную части собственного ядра терминальной полоски. В медиальную группу включается также сошниково-носовая миндалина или та часть миндалины, которая связана с добавочной обонятельной системой. Эта подгруппа заключает в себе массу медиального миндалевидного ядра, собственное ядро добавочного обонятельного тракта и каудомедиальное корковое миндалевидное ядро. Переходное поле миндалина гиппокамп на основании своих связей также включается в медиальный миндалевидный комплекс. Базально-латеральная миндалевидная группа включает латеральное и базальнолатеральное миндалевидные ядра и часть базально-медиального ядра.
Центральная миндалевидная группа включает центральное миндалевидное ядро и тесно связанные с ним дорсолатеральные части подчечевицеподобного безымянного вещества, а также латеральную и вентральную части собственного ядра терминальной полоски. Наконец, базальномедиальное ядро может быть отнесено, частично, к медиальному миндалевидному комплексу или к базально-латеральной миндалевидной группе.
Базальные ядра — nucll. basales, часто именуются базальными ганглиями, или подкорковыми ядрами; расположены в основании полушарий большого мозга. Одним из главных составляющих базальных ядер является полосатое тело, образованное комплексом хвостатое тело-скорлупа — nucl. caudatus-putainen и бледным шаром — pallidum (globus pallidus). Два других тесно связанных ядра - субталамическое ядро и черное вещество — часто рассматриваются вместе с полосатым телом. В состав базальных ядер включаются также ограда и миндалевидное тело.
Термин «комплекс хвостатое ядро-скорлупа» обычно относится к полосатому телу — corp. striatum (или неостриатуму — neostriatum), и это обозначение особенно уместно у крысы, где оно формирует крупную «полосатую» массу, пронизанную рассеянными пучками волокон, представляющих кортикофугальные (идущие от коры) и кортикопетальные (идущие к коре) связи.
Островки Каллеха — insulae Calleja — являются заметными образованиями в нескольких областях базального переднего мозга, включая вентральную область полосатого тела и миндалевидное тело. Представляют собой отдельные скопления, или островки, плотноупакованных клеток мелкого или среднего размеров. Основная протяженность островков отмечена в рострокаудальном направлении. Самый крупный островок — большой островок — insula magna — всегда расположен вдоль медиального края nucl. accumbens.
Ограда — claustrum — представляет собой длинную пластинку серого вещества, расположенную латеральнее комплекса хвостатое ядро-скорлупа. Рострально имеет наибольший размер, форму широкого полумесяца. Медиально граничит с малым щипцом мозолистого тела и расположена дорсальнее nucl. accumbens.
Среди систем волокон переднего мозга выделяют пояс — cingulum, внутреннюю капсулу — capsula int., ассоциативные и комиссуральные проводящие пути, включающие ростральную спайку — commissure rostralis, спайки гиппокампа и мозолистое тело, а также шесть основных поперечных проводящих путей: свод, вентральную ножку таламуса, мозговую полоску, сосцевидно-таламический тракт, отогнутый пучок и терминальную полоску — stria terminalis (пять первых путей описаны выше).
Мозолистое тело — corp. callosum — является крупнейшим волокнистым образованием, соединяющим новую кору обоих полушарий; представляет собой участок белого вещества головного мозга, расположенный в глубине продольной щели между полушариями большого мозга. В мозолистом теле различают ствол мозолистого тела — truncus corporis callosi, каудальную часть тела — валик мозолистого тела — splenium corporis callosi, ростральную часть — колено мозолистого тела — genu corporis callosi. Кроме того, выделяют покров мозолистого тела — tapetuin corporis callosi, являющийся соединительными волокнами на стороне бокового желудочка, и лучистость мозолистого тела — radiatio corporis callosi, состоящую из больших щипцов — forceps major — и малых щипцов — forceps minor.
Боковой желудочек (I и II) — ventnculus lat. — парный желудочек головного мозга, находящийся в полушариях большого мозга. Боковые желудочки образуют пару цилиндров, размещенных в левом и правом полушариях; сообщаются с III желудочком посредством межжелудочкового, или монроева, отверстия — for. interventriculare s. Monro. В отличие от человека, у крысы височный (нижний) рог — cornu temporale — недоразвит, а задний рог — практически отсутствует. За вытянутым ростральным (передним) рогом — cornu rostrale (ant.) — идет относительно большая центральная часть — pars centralis — желудочка, продолжающаяся в височный рог; последний заканчивается на уровне каудальной части миндалевидного тела.
Мозговые оболочки
Мозговые оболочки — meninges — представляют собой три непрерывные соединительнотканные мембраны, окружающие и защищающие центральную нервную систему. Различают твердую, паутинную и мягкую оболочки головного и спинного мозга.
Твердая оболочка головного мозга — dura mater encephali, иногда носит название пахименинкс — pachymeninx, является наружной оболочкой, состоящей из плотной волокнистой соединительной ткани, которая защищает мозг. Прилежит в полости черепа к внутренней поверхности костей. Между надкостницей и твердой оболочкой располагаются венозные синусы. Твердая оболочка своей наружной пластинкой образует влагалище для черепных нервов, а внутренней пластинкой покрывает поверхность полушарий головного мозга, углубляясь в продольную щель большого мозга в виде серпа большого мозга — falx cerebri. Соединительнотканная стенка, начинающаяся от твердой оболочки и отделяющая оба полушария от мозжечка, называется намётом мозжечка — tentorium cerebelli. Между твердой и паутинной оболочками располагается субдуральное пространство — cavum subdurale, почти неразличимое в прижизненных препаратах.
Твердая оболочка спинного мозга — dura mater spinalis — плотная, фиброзная, защищает спинной мозг. Состоит из двух листков — наружного, сливающегося с надкостницей позвонков, и внутреннего — собственно оболочки спинного мозга. С мозга она переходит на спинномозговые нервы в местах их отхождения и образует влагалища, в которых на небольшом участке идут нервы. Между твердой оболочкой и надкостницей позвоночного канала образуется полость — эпидуральное пространство — cavum epidurale. Нитеобразный каудальный конец твердой оболочки называется нитью твердой оболочки спинного мозга — filum durae matris spinale.
Паутинная оболочка головного мозга — arachnoidea encephali — является частью лептоменинкса — leptoineninx — мягкой оболочки головного мозга, состоящей из паутинной и сосудистой оболочек. Паутинная оболочка очень тонкая, нежная, не имеет сосудов, состоит из рыхлой волокнистой соединительной ткани и связана с мягкой оболочкой головного мозга соединительнотканными трабекулами (перекладинами) — trabeculae, образующими систему ячеек под паутинного пространства — cavum subarachnoideale, заполненную спинномозговой жидкостью.
На основании мозга и при переходе его в спинной мозг образуются расширения подпаутинной полости — подпаутинные цистерны -cisternae subarachnoideales. В синусы твердой мозговой оболочки в виде узелков выпячиваются ворсинки паутинной оболочки — грануляции паутинной оболочки (пахионовы грануляции) — granulationes arachnoideales (— Pachioni — BNA; granula meningica — JNA).
Паутинная оболочка спинного мозга — arachnoidea spinalis — является продолжением паутинной оболочки головного мозга, расположена между твердой и мягкой оболочками спинного мозга. От твердой оболочки отделена субдуральным пространством, от мягкой оболочки — подпаутинным пространством. Пространство спинного мозга соединяется с аналогичным пространством головного мозга и заполнено спинномозговой жидкостью.
Мягкая (сосудистая) оболочка головного мозга — pia mater encephali — прилежит непосредственно к веществу головного мозга и повторяет рельеф его поверхности. Она проникает также и в полости мозга и образует сосудистую основу — tela choroidea (chorioidea) — и сосудистое сплетение — pi. choroideus (chorioideus) — каждого из четырех желудочков мозга (подробнее см. при описании каждого из желудочков). Мягкая оболочка состоит из рыхлой волокнистой соединительной ткани с большим количеством кровеносных сосудов, питающих головной мозг.
Мягкая (сосудистая) оболочка спинного мозга — pia mater spinalis — является внутренним слоем лептоменинкса, покрывает спинной мозг и проникает в мозговое вещество, пронизана кровеносными сосудами. На боковых сторонах спинного мозга оболочка образует множественные фиброзные зацепки — зубчатые связки — ligg. denticulati, которые прикрепляются справа и слева своими основаниями к мягкой оболочке, вершинами — к твердой мозговой облочке и служат для укрепления спинного мозга.
Скачать реферат:
Пароль на архив: privetstudent.com