Эндокринные, или железы внутренней секреции — все железы или группы клеток, продукты которых, гормоны или секреты, из-за отсутствия собственных выводящих путей выделяются в кровеносные и лимфатические капилляры и распространяются по всему телу через систему кровообращения. При контакте гормонов, поступающих в органы, лежащие около, а чаще далеко от места выработки гормонов, со специфическими рецепторами, они оказывают тормозящее или активирующее действие, и часто с вегетативной нервной системой, на органы, принимающие участие в обмене веществ и морфологические изменения. Это способствует адекватному приспособлению органов, принимающих участие в обмене веществ, к условиям окружающей среды. В отличие от гормонов, паракринные сигнальные вещества при диффузии в и интерстициальной ткани оказывают влияние на клетки или группы клеток, которые располагаются около места выработки продуктов.
Далее будут подробно рассмотрены только макроскопически различимые железы, вырабатывающие гормоны, параганглии, а также островки поджелудочной железы. Так, в стенке желудка и кишечника располагаются многочисленные, отдельно лежащие клетки, которые, несмотря на различия в строении и вырабатываемых продуктах, объединяют в энтероэндокрин-ную систему. Сходные по строению клетки располагаются в слизистой оболочке бронхов и уретры, а также в почках (Andrew, 1981; Bohme, 1992; Grube, 1986; Hanyu et al., 1987; Kitamura et al., 1982; Pearse, 1980). В миокарде располагаются клетки, которые за счет атриального натриурети-ческого пептида (АНП) при выработке натрия в почках, оказывают непрямое влияние на объём экстрацеллюлярной жидкости (Forssmann, 1987).
Насколько тесно осуществляется взаимодействие эндокринных органов и вегетативной нервной системы, которое можно рассматривать как функциональное единство при регуляции процессов, происходящих в организме, можно уяснить из следующего: 1) в центральной нервной системе происходит интенсивное взаимодействие ядер промежуточного мозга с гипофизом и шишковидной железой, 2) как клетки энтероэндокринной системы, так и вегетативной нервной системы продуцируют и выделяют нейропептиды.
ГИПОФИЗ
Гипофиз, hypophysis, glandula pituitaria, представляет собой непарный небольшой орган, расположенный между chiasma opticum и corpus mamillare вентрально к промежуточному мозгу. Он состоит из формирующегося на базе промежуточного мозга нейрогипофиза, и из аденогипофиза, возникающего из гипофизарного кармана выстилки крыши ротовой полости. В нейрогипофизе различают воронку, infundibulum, или ножку гипофиза, и lobus nervosus, или задняя доля (-/2). Аденогипофиз включает pars tuberalis, или воронокообразную долю (-/3), pars distalis, или переднюю долю (-/3'), pars intermedia, или промежуточную долю (-/4). Гипофиз является составной частью гипоталамусо-гипофизарной системы. Это выражается в том, что гормоны, выделяющиеся в кровь в нейрогипофизе, образуются нейросекреторными нейронами, тела которых находятся в nucleus supraopticus и nucleus paraventricularis гипоталамуса. А функционирование аденогипофиза управляется либеринами и статинами, которые выделяют нейроны мелкоклеточных ядер серого бугра, tuber cinereum.
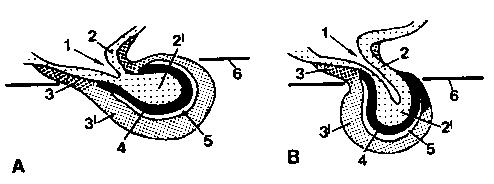
Рис. 1. Схематическое изображение гипофиза по средней линии собаки (А) и кошки (В)
1 recessus infundibuli; 2 infundibulum, 2' lobus nervosus neurohypophysis; 3 pars tuberalis, 3' pars distalis adenohypophysis; 4 pars intermedia adenohypophysis; 5 cavum hypophysis; 6 dura mater
Гипофиз у собак несколько сплющен, овальный, у кошек шарообразный. Размер гипофиза не только зависит от породы, но даже в пределах одной породы имеются индивидуальные различия (Latimer, 1942, 1965; White/Foust, 1944; Hanstrom, 1966). Размер гипофиза собаки со средним размером головы составляет 10 х 7 х 5 мм, кошки — 5 х 5 х 2 мм. При одинаковых условиях содержания у самок гипофиз несколько крупнее, чем у самцов, у беременных животных крупнее и тяжелее, чем у небеременных (Latimer, 1942; White/ Foust, 1944). Масса гипофиза кобелей различных пород со средней массой тела в 11 кг составляет 0, 0658 г, сук со средней массой тела в 8, 93 кг — 0, 067 г (Latimer, 1942).
Нейрогипофиз, neurohypophysis, посредством ножки или воронки гипофиза, infundibulum, находится в непосредственной связи с tuber cinereum гипоталамуса. Ножка гипофиза цилиндрическая, очень короткая и содержит в виде углубления в проксимальной части короткий, а у кошек, достигающий lobus nervosus, recessus infundibuli (-/1). Дистально ножка гипофиза толще и проходит без четкой границы в lobus nervosus, или заднюю долю (-/2’).
Аденогипофиз, adenohypophysis, крупнее, чем нейрогипофиз. Его pars tuberalis, тубералъная или воронкообразная часть, охватывает у собак и кошек в ножку гипофиза. У собак также передняя и промежуточная доли аденогипофиза (-/3', 4) охватывают заднюю долю нейрогипофиза со всех сторон, в то время как у кошек проксимальный участок каудальной поверхности задней доли остается непокрытым. При развитии между передней и промежуточной долями аденогипофиза у собак и кошек остается гипофизарная полость, cavum hypophysis (-/ 5), которая в значительной степени варьирует по своей длине и ширине.
В свежем органе поверхность разреза нейрогипофиза выглядит из-за большого количества нейритов и глиальных клеток гомогенной и стекловидной, поверхность разреза аденогипофиза, в которой преобладают эпителиальные клетки и синусоидные капилляры, зернистая консистенция плотнее, чем у нейрогипофиза. Особенности микроскопического строения гипофиза, а также роль различных типов клеток при выработке отдельных гормонов, а также влияние на другие железы, выделяющие гормоны, или другие органы, описаны в учебниках по гистологии и физиологии (напр., Mosimann/Kohler, 1990; Scheunert/Trautmann, 1987).
К задней доле гипофиза непосредственно идут только аа. hypophysiales caudales. Они возникают у собак из каудальной соединительной ветви а. intercarotica caudaiis, которая проходит в твердой оболочке по телу базисфеноида. У кошек эти сосуды идут от rete mirabile epidurale. После прохождения a. carotis interna через диафрагму седла, diaphragma sellae от нее, или от отходящей от нее а. cerebri rostralis, отделяются аа. hypophysiales rostrales, которые идут к ножке гипофиза и задней доле аденогипофиза. Часто мелкие аа. hypophysiales rostrales возникают с каждой стороны каудальной соединительной артерии, a. communicans caudaiis, и идут радиально, сходясь на ножке гипофиза. В твердой оболочке мозга у гипофиза артерии гипофиза соединяются в одну тонкую сеть, сплетение (Green, 1951), от которого артерии, прежде всего, идут к срединному возвышению, eminentia mediana и infundibulum нейрогипофиза, а также к pars tuberalis аденогипофиза. Из этой первичной капиллярной области в ножке гипофиза формируются многочисленные вены, которые идут дистально по вентральной поверхности аденогипофиза, а далее — в объемистые синусоиды передней и промежуточной долей. Эта система делает возможным влияние ли-беринов и статинов, вырабатываемых в tuber cinereum, а далее перемещающихся по tractus tuberoinfumdibularis в ножку гипофиза, после их дальнейшего транспорта в крови, на различные клетки передней доли. Многочисленные вены, которые отводят кровь от гипофиза, впадают вскоре в sinus cavernosus или каудально лежащий sinus intercavemosus.
Симпатические нервные волокна из краниального шейного ганглия идут к гипофизу либо в виде периваскулярного сплетения с аа. hypophysiales или в виде ветвей n. caroticus internus.
На наружной поверхности гипофиза твердая оболочка мозга формирует тонкую соединительнотканную капсулу, которая одновременно представляет собой прочное соединение гипофиза с плоской ямкой гипофиза, fossa hypophysiales, на теле ба-зисфеноида. В области ножки гипофиза твердая мозговая оболочка выдается над свободным краем турецкого седла, sella turcica в видediaphragma sellae, покрывает большую часть гипофиза с дорсальной стороны и оставляет только небольшое отверстие для прохождения ножки гипофиза. В этой области заканчивается, по отношению к гипофизу, и cavum subarachnoidale, которая особенно обширна с дорсальной стороны от него в виде межножковой цистерны, cisterna interpeduncularis. Между двумя пластинками твердой оболочки по обеим сторонам гипофиза проходит sinus cavernosus, а каудально от него sinus intercavernosus. В области последнего с каждой стороны к гипофизу идут a. carotis interna, или соответственно, у кошек — rete mirabile epidurale, n. oculomotorius, n. trochlearis и n. ophthalmicus, a также n. abducens.
ШИШКОВИДНАЯ ЖЕЛЕЗА (ЭПИФИЗ)
Шишковидная железа, glandula pinealis, представляет собой непарный орган. Его поперечное сечение круглое. Шишковидная железа лежит между большими полушариями впереди крыши среднего мозга, tectum mesencephali. Его размер варьирует у разных животных, и у собак среднего размера длина достигает примерно 3 мм, а диаметр — 2 мм. У кошек это соотношение составляет 2x1 мм. Являясь частью промежуточного мозга, шишковидная железа соединяется с каудальным участком его крыши посредством уздечек, habenulae с короткой ножкой, pedunculus. Через это соединение проходят волокна спайки уздечек, comissura habenularum. В теле, corpus, шишковидной железы, кроме нервных волокон, находятся пинеалоциты, которые в зависимости от продолжительности и интенсивности света вырабатывают гормон мелатонин. У собак и у кошек, вне зависимости от возраста, особенно на вентральной поверхности шишковидной железы, некоторые пинеалоциты содержат меланин. Функциональное значение этих пигментированных клеток еще не изучено (Calvo et al., 1992). Учитывая соединение с промежуточным мозгом, а также гуморальное взаимодействие с другими железами, выделяющими гормоны, посредством мелатонина шишковидная железа представляет важный центральный орган нейровегетативной регуляции. Ночью выработка мелатонина происходит активнее, чем днем, и при участии обратной связи через краниальный шейный ганглий симпатической части нервной системы и при иннервации симпатическими волокнами шишковидная железа может оказывать контролирующее влияние на биологические ритмы. Артерии, которые около шишковидной железы снабжают pia mater, посылают тонкие ветви во внутреннюю часть органа. В шишковидной железе ветви разветвляются в синусоиды.
ЩИТОВИДНАЯ железа
Щитовидная железа, glandula thyreoidea, состоит из левой и правой долей, lobus sinister
A) et lobus dexter, а также соединяющего их перешейка, isthmus. Форма каждой доли у собак и кошек значительно варьирует, является овальной, а по бокам немного сплющенной, и у кошек, чаще всего, тоньше, чем у собак. Доли, от темно-красно-коричне-вого до серо-красного цвета имеют консистенцию, сходную с консистенцией печени. У взрослых животных щитовидная железа может быть плотнее, а у кошек — более мягкой. Частота возникновения перешейка у кошек различна (16-87%). У собак зависит от размера тела. Перешеек имеется у половины крупных собак, у трети собак среднего размера и четверти мелких собак (Heller, 1932). Обе доли располагаются у собак на дорсолатеральной поверхности трахеи и идут параллельно ей. В редких случаях железа может располагаться немного краниально или каудально. При ультразвуковом исследовании у собак щитовидная железа выявляется каудально от гортани в виде гомогенной веретенообразной структуры и четко ограничена от окружающих структур (Wisner et al., 1991). У кошек обе доли выше с дорсальной стороны, чем у собак, поэтому может располагаться между трахеей и пищеводом и дорсолатерально быть покрыта m. longus capitis. При наличии перешейка соединяются каудальные полюса обеих долей, и перешеек проходит по вентральной поверхности трахеи. Эпителиальные клетки фолликулов щитовидной железы продуцируют гормоны тироксин и трийодтиронин, играющие большую роль в процессах обмена веществ. Эти эпителиальные клетки в процессе развития отделяются от эпителия корня языка. Затем они через ductus thyreoglossus достигают боковых поверхностей первого кольца трахеи. Между этими клетками всегда располагаются так называемые С-клетки. Они продуцируют кальцитонин, который вместе с паратгормоном участвует в поддержании постоянного содержания кальция.
Относительная масса щитовидной железы у собак и кошек ко времени рождения максимальна, а в первые недели после рождения уменьшается. Независимо от породы абсолютная и относительная масса щитовидной железы варьируют.
Абсолютная и относительная масса щитовидной железы у собак и кошек
(Haensly et al., 1964; Heller, 1932; Latimer, 1939; Meissner, 1924; Meyer, 1952; Schneebeli, 1958; Schweinhuber, 1910):
Таблица 1
| Абсолютный вес в мг |
Относительная масса, в мг/кг массы тела |
|
| Взрослая собака | (115) 560-9770(-25300) | (15-) 84—342 |
| Щенок | 25—222 | 210—360 |
| Взрослая кошка | 35—1950 (-7200) | 80—120 |
| Котенок | 20—40 | 135—200 |
Добавочные щитовидные железы, glandulae thyroideae accessoriae, могут формироваться в процессе развития от отделившихся частей щитовидной железы, что у собак встречается чаще, чем у кошек. Они могут встречаться в основании языка, вдоль шеи, в средостении около сердца или около дуги аорты. Их размер очень варьирует, и они часто могут быть выявлены только при гистологическом исследовании. Если при развитии сохраняется часть ductus thyreoglossus, он может развиться до кисты в области шеи.
Основным сосудом, снабжающим щитовидную железу, является a. thyreoidea cranialis. Она возникает из a. carotis communis на уровне кольцевиднотрахейной мембраны (связки), membrana cricotrachlealis или первого трахейного хряща. Кроме ветвей к глотке, гортани и прилегающей мускулатуре эта артерия отдает ramus dorsalis et ramus ventralis вдоль соответствующих частей каждой доли щитовидной железы как к щитовидной железе, так и к эпителиальным тельцам. Область возникновения тонкой а. thyreoidea caudalis (-/1) варьирует. Чаще всего, она возникает из плечеголовного ствола, truncus brachiocephalicus или реберно-шейного ствола, truncus costocervicalis. Реже она формируется от правой подключичной артерии, a. subclavia dextra. A. thyreoidea caudalis всегда сопровождает n. laryngeus recurrens (-/5) и соединяется через анастомозы с дорсальной ветвью a. thyreoidea cranialis.
Экстрагландулярные вены различны не только у разных животных, но и на различных сторонах тела одного и того же животного, и соединяются друг с другом. V. thyreoidea cranialis (-/ п) и часто двойная v. thyreoidea media (-/t) отводит кровь в v. jugularis на своей стороне. Arcus laryngeus caudalis (-/р) представляет собой соединение между левой и правой v. thyreoidea cranialis, а также краниальной частью непарной v. thyreoidea caudalis (-/u). Последний сосуд проходит по средней линии по вентральной поверхности трахеи и впадает либо в левую, либо правую v. brachiocephalica или v. jugularis externa, или interna правой стороны.
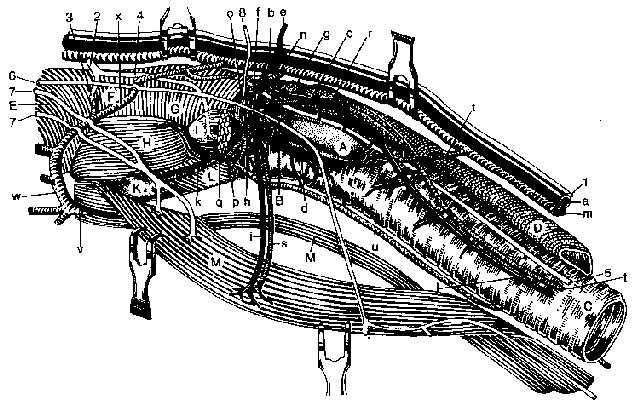
Рис. 2. Топография щитовидной железы и левых наружных эпителиальных телец собаки (по Borer, 1990)
A glandula thyreoidea; В glandula parathyreoidea; С trachea; D oesophagus; E m. hyopharyngeus; F m. thyreopharyngeus; G m. cricofaringeus; H m. thyreohyoideus; L m. sternothyreoideus; К cartilago thyreoidea; L m. cricothyreoideus; M m. sternohyoideus
a a. carotis communis; b a. thyreoidea cranialis; c — k ветви a. thyreoidea cranialis; с ramus dorsalis; d ramus ventralis; e ramus sternoclei domastoideus; f ramus laryngeus caudalis; g ramus pharyngeus; h ramus cricothyreoideus; i ramus muscularis; k ramus laryngeus; I a. thyreoidea caudalis; m v. jugularis interna; n v. thyreoidea cranialis; o — s aste der v. thyreoidea cranialis; t v. thyreoidea media; u v. thyreoidea caudalis; v v. laryngea impar; w arcus hyoideus; x anastomose zwischen arcus hyoideus und v. jugularis interna
1 truncus vagosympathicus; 2 n. laryngeus cranialis; 3 ramus internus n. laryngeus cranialis; 4 ramus externus n. laryngeus; 5 n. laryngeus recurrens; 6, 7 rami musculares из ansa cervicalis; 8 соединительная ветвь к 1-му шейному нерву
В щитовидной железе лимфатические капилляры формируют вокруг отдельных фолликулов плотную сеть (Rusznyak et al., 1967), и отводящие лимфатические сосуды идут к In. retropharyngeus medialis.
Симпатические нервы к щитовидной железе формируются из краниального шейного ганглия, а парасимпатические — из n. laryngeus cranialis. Отдельные волокна могут выходить из окончания n. laryngeus recurrens.
ЭПИТЕЛИАЛЬНЫЕ ТЕЛЬЦА (ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ)
После развития из эпителия третьего и четвертого жаберного кармана наружное эпителиальное тельце, glandula parathyreoidea externa, также называется glandula parathyreoidea IV и внутреннее, glandula parathyreoidea interna, также называемое glandula parathyreoidea III. Вырабатываемый ими паратгормон, вместе с кальцитонином, секретируемым С-клетками щитовидной железы, регулирует обмен кальция.
У собак наружное эпителиальное тельце имеет чечевицеобразную форму или форму рисового зерна с гладкой поверхностью и располагается у краниального полюса или краниальной половины доли щитовидной железы, реже — около дорсального края. У кошек наружное эпителиальное тельце, как правило, располагается латераль-но в каудальной половине доли щитовидной железы. Размер и масса эпителиальных телец у собак до года мало зависят от возраста. Размер наружного эпителиального тельца у крупных собак составляет 3-7 х 2-5, 5 х 1, 5-2, 5 мм, внутреннее эпителиальное тельце немного меньше. Цвет варьирует от золотисто-желтого цвета до красновато-коричневого и часто хорошо выделяется на фоне щитовидной железы.
Внутреннее эпителиальное тельце лежит у собак и у кошек в средней части долей щитовидной железы в паренхиме щитовидной железы, несколько удаленное от медиальной или дорсальной поверхности и не всегда заметное снаружи. В некоторых случаях оно может и отсутствовать (Pinto е Silva, 1947).
Наружное эпителиальное тельце получает 1-2 rami glandulares от a. thyreoidea cranialis, а венозный отток осуществляется через rami glandulares, которые впадают в v. thyreoidea cranialis или arcus laryngeus caudalis. Внутреннее эпителиальное тельце не имеет собственных артериальных или венозных ветвей, но примыкает к сосудам щитовидной железы (Orsi et al., 1975).
Симпатические волокна из краниального шейного ганглия достигают эпителиальные тельца, сопровождая артерии, парасимпатические волокна происходят из n. laryngeus recurrens.
НАДПОЧЕЧНИКИ
Надпочечник, glandula suprarenalis или adrenalis, — парный орган, который состоит из коркового вещества, cortex и мозгового вещества, medulla (-/С, 2). Снаружи этот орган имеет тонкую соединительнотканную капсулу и окружен соединительной тканью с жировыми клетками, располагаясь с медиальной стороны от краниального полюса почки ретропе-ритонеально. Кора надпочечников развивается из мезодермы и на нее оказывает влияние, в основном, АКТГ аденогипофиза. Мозговое вещество надпочечников продуцирует адреналин и норадреналин и, прежде всего, регулируется симпатической частью нервной системы. На поверхности разреза свежего надпочечника граница между светлой корой и темным мозговым веществом хорошо заметна макроскопически. У собак каждый надпочечник (-/А) удлинен, дор-совентрально уплощен и имеет цвет от светлосерого до белого. У кошек желтовато-белые надпочечники (-/В) короче, чем у собак, овальные и дискообразные. По вентральной поверхности проходит общий ствол каудальной диафрагмальной вены, v. phrenica caudaiis, и краниальная брюшная вена, v. abdominalis cranialis, оставляя у кошек поверхностную, а у собак глубокую борозду. Из-за наличия этой глубокой борозды у собак на правом надпочечннке могут выделяться две не полностью отделенные друг от друга удлиненные, а на правом надпочечнике две округлые доли. Общий ствол a. phrenica caudaiis и a. abdominalis cranialis проходит надпочечники с дорсальной стороны и не оставляет борозды.
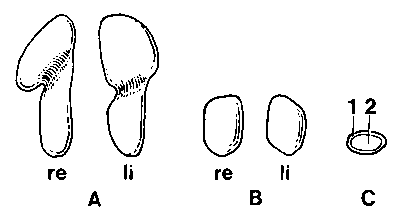
Рис. 3. Надпочечники собаки (А) и кошки (В), вид с вентральной стороны; С — поперечное сечение 1 cortex; 2 medulla. Натуральная величина
У взрослых самок, а также беременных и лактирующих, размер и масса больше, чем у самцов, а также молодых животных.
У собак и кошек надпочечники располагаются ретроперитонеальио и медиально от краниальной половины почек, или медиально от их краниального полюса. Левый надпочечник соединен с левой стенкой aorta caudaiis, правый с правой стенкой v. cava caudaiis.
Кровоснабжение надпочечников осуществляется многочисленными аа. suprarenales или непосредственно из aorta abdominalis, или из a. phrenica caudaiis, a. abdominalis cranialis или a. renalis. После прохождения через соединительнотканную капсулу эти ветви разветвляются и отдают радиально объемистые капилляры но всей окружности к мозговому веществу. Из капиллярной сети мозгового вещества кровь собирается в крупную центральную вену и далее идет по нескольким vv. suprarenales в v. cava caudalis, v. phrenica caudalis, v. abdominalis cranialis или v. renalis. Имеются значительные индивидуальные различия в количестве, длине кровеносных сосудов. У собак и кошек мельчайшие сосуды объединяются в соединительной ткани между почками и надпочечниками, что, вероятно, объясняет то, что, по крайней мере, небольшая часть продуцируемых в мозговом веществе надпочечников катехоламинов может попадать в почки кратчайшим путем (Christe, 1980; Dempster, 1978; Earle/Gilmore, 1982). Лимфатические капилляры многочисленны во всех частях надпочечников и располагаются в виде сети. По нескольким лимфатическим сосудам лимфа собирается в Inn. lumbale aortic.
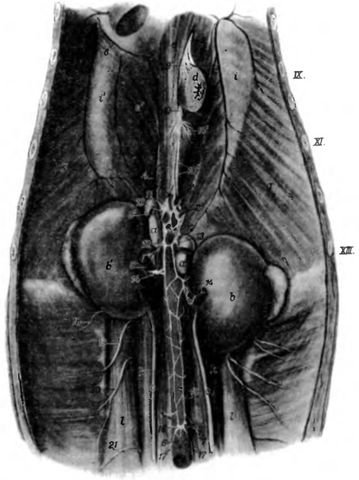
Рис. 4. Расположение надпочечников таксы с расположенными рядом ганглиями и нерками (по Seiferle, 1992) а левый, а’ правый надпочечник; b левая, b' правая почка; с мочеточник; d oesophagus; e вентральная ножка, е’ боковая ножка правой части диафрагмы. f левая часть диафрагмы; g pars costalis мускулатуры диафрагмы; h v cava caudaiis; i. i' зеркало диафрагмы; k m. psoas minor; l m. psoas major; IX. —XIII. 9— 13. ребра
1 aorta abdominalis; 2 a. hepatica, 7 a. gastrica sinistra, 2" селезеночная артерия a. coeliaca;3 a. mesenterica cranialis 4 a. phrenica caudaiis; 5 a. und v. renalis, 6 a. mesenterica caudaiis; 7 a. tcsticularis; 8 vv. phrenicac, 8' общий ствол v. phrenica caudaiis и v. abdominalis cranialis; 9 truncus vagalis ventralis, 9' его rami gastrici parietales; 10 truncus vagalis dorsalis, 10' его rami gastrici visccrales, 10" его rami cocliaci; 11 ganglion cocliacum; 12ganglion mesentericum craniale; 13 ветви plexus suprarenalis; 14 ganglion renale и plexus renalis; 15 plexus aorticus abdominalis; 16 ganglion mesentericum caudale; 17 левый и правый n. hypogastricus; 18 ветвь n. iliohvpogastricus cranialis; 19 ветвь n iliohypogastricus caudaiis; 20 n. ilioinguinalis; 21 n. cutaneus femoris lateralis
Таблица 2
|
|
Длина |
Ширина |
Толщина |
Масса |
|
в см |
в см |
в см |
в мг |
|
|
Собака |
0, 8—3, 3 |
0. 6—1. 8 |
0. 2—0, 8 |
60 -1600 |
|
Кошка |
0. 8—1, 5 |
0. 5—1. 0 |
0, 2—0. 4 |
40—400 |
Многочисленные вегетативные нервные волокна к надпочечникам возникают либо непосредственно из располагающихся рядом n. splanchnicus major, либо из ganglion coeliacum и ganglion mesentericum craniale. В виде plexus suprarenalis они достигают надпочечники непосредственно или с кровеносными сосудами и входят с ними в орган. Нервные волокна формируют в капсуле сплетение, от которого отходят многочисленные пучки нервных волокон в кору и мозговое вещество надпочечников.
островки поджелудочной ЖЕЛЕЗЫ
Внутри поджелудочной железы между экзок-ринными клетками концевых отделов железы в небольшие группы, островки поджелудочной железы или островки Лангерганса, insulae pancreaticae, объединяются эндокринные клетки поджелудочной железы, endocrinocyti pancreatici. Отдельные, включающие большое количество сосудов островки неравного размера, состоят из 10-100 эндокриноци-тов. Число островков значительно варьирует у собак и кошек, достигает нескольких тысяч. В lobus sinister поджелудочной железы островки Лангерганса крупнее и многочисленнее, чем в lobus dexter. Капиллярная область инкреторных и внешнесекреторных частей поджелудочной железы соединяются друг с другом, причем просвет капилляров в островках больше и капилляры многочисленнее, чем в экзокринной части поджелудочной железы. Переход к пограничной поверхности между двумя частями очень выражен.
При микроскопическом исследовании в островках выделяют 3 типа клеток. А-клетки составляют в целом 10 — 20% эндокринных клеток, однако отсутствуют в островках каудальной части lobus dexter поджелудочной железы. Это может быть связано с тем, что каудальная часть lobus dexter и остальные части имеют различное происхождение. А- клетки продуцируют глюкатон и регулируют с ин-сулинпродуцирующими В-клетками обмен углеводов. В-клетки составляют 80-90% клеток эндокринной железы. Кроме продуцирующих соматостатин D-клеток, которые составляют 1% от всех клеток островка, также имеются и другие, отдельно встречающиеся, клетки, которые, например, могут вырабатывать гастрин и серотонин. Эти клетки сравниваются с клетками энтероэндокринной системы (Mosimann/Kohler, 1990). Электронная микроскопия выявила наличие в А-клетках электронноплотных гранул диаметром до 0, 5 μт. В В-клетках гранулы крупнее, чем в А-клетках, обладают меньшей электронной плотностью, однако содержат кристаллические включения. В D-клетках гранулы мельче и обладают меньшей электронной плотностью, чем гранулы А-клеток.
ПАРАГАНГЛИИ
Точного определения того, что такое параганглии, не существует. Чаще всего, параганглиями называют крупные или мелкие скопления лишенных отростков, катехоламинсодержащих, хромаффинных клеток, которые располагаются в непосредственной близости от ганглиев автономной нервной системы или от больших артерий. Чаще всего, эти скопления различаются только с помощью макро-мик-роскопических методов исследования. Так как эти клетки, а также клетки мозгового вещества надпочечников имеют общее происхождение, долгое время считалось, что клетки параганглиев обладают инкреторной активностью. Сегодня известно, что мозговое вещество надпочечников, как наиболее крупный параганглий, согласно данному определению активно продуцирует гормоны, но сонный клубок, glomus caroticum, а также аортальный клубок, glomus aorticum функционируют как хеморецепторы и регистрируют парциальное давление СO2 в крови.
Glomus caroticum у собак имеют очень тонкую капсулу из рыхлой соединительной ткани, которая без четкой границы переходит в окружающие ткани. Поэтому границы между glomus и окружающими тканями мало заметны при рассмотрении через лупу. Располагается, чаще всего, краниомедиально от конечного деления a. carotis communis в области возникновения а. pharyngea ascendens или a. occipitalis, реже — в области возникновения a. carotis interna. Glomus caroticum шарообразной формы или удлинен, иногда охватывает наподобие кольца или полукольца область возникновения одной из названных артерий (Cantieni/Frewein, 1982). Точные данные по размеpy glomus caroticum могут быть получены при гистоморфометрических исследованиях. Объем glomus caroticum у взрослой немецкой овчарки и взрослого боксера составляет 3-16 мм3. Густая сеть объемистых капилляров контактирует с паренхиматозными клетками (Тип I и Тип II). В среднем, у собак имеется 3, 3% клеток I Типа, 2, 2% клеток II типа (Frei-Kuchen, 1981; Pallot, 1987).
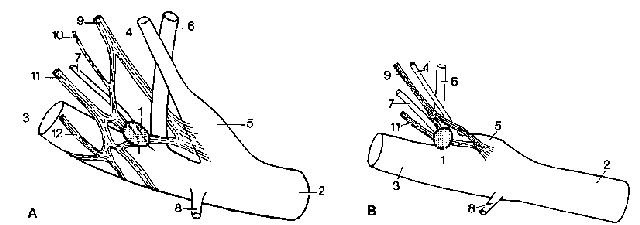
Рис. 5. Топография правого glomus caroticum, вид с медиальной стороны. А — немецкой овчарки (по Cantieni/ Frewein, 1982) и В — кошки (изменено, по Pallot, 1987)
1 glomus caroticum; 2 a. carotis communis; 3 a. carotis externa; 4 a. carotis interna; 5 sinus carotis; 6 a. occipitalis; 7 a. pharyngea ascendens; 8 a. larvngea cranialis; 9 ramus sinus carotici языкоглоточного нерва; 10 ветвь n. vagus; 11 ветвь от ganglion cervicale craniale; 12 plexus caroticus externus
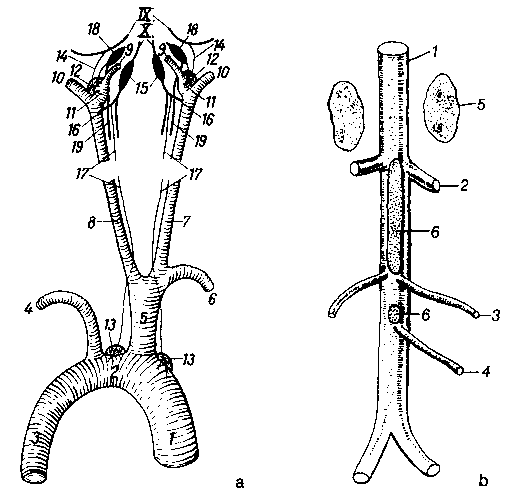
Рис. 6. а. Схематическое изображение параганглиев головы, шеи и грудной области (по Seiferle, 1992)
1 aorta descensens; 2 arcus aortae; aorta thoracica; 4 a. subclavia sinistra; 5 truncus brachiocephalicus; 6 a. subclavia dextra; 7 a. carotis communis dextra; 8 a. carotis communis sinistra; 9 a. carotis interna; 10 a. carotis externa;
11 sinus caroticus; 12 glomus caroticum; 13 glomus aorticum; 14 ramus sinus carotici; 15 ganglion distale блуждающего нерва; 16 n. laryngeus cranialis; 17 n. depressor; 18 ganglion cervicale craniale; 19 симпатическая часть
IX n. glossopharyngeus; X n. vagus
Рис. 6. b. Схематическое изображение крупных абдоминальных параганглиев собаки в возрасте 24 недель. Вид с вентральной стороны (по Mascorro/Yates, из Seiferle/Bohme, 1992)
1 aorta abdominalis; 2 a. renalis; 3 a. testicularis (ovarica); 4 a. mesenterica caudalis; 5 надпочечник; 6 paraganglion aorticum abdominale
У кошек glomus caroticum из-за мощной соединительнотканной капсулы отделяется от окружающих тканей легче, чем у собак. В целом, glomus caroticum шарообразный диаметр, составляет 2 мм и располагается на месте возникновения либо a. pharyngea ascendens, либо а. occipitalis. Составные части, по Seiferleet al. (1977), включают: сосуды 22, 3%, специфическая ткань 16, 9% и остальная ткань 60, 8%.
У собак и кошек glomus caroticum иннервируется ответвлениями от ramus sinus carotici языкоглоточного нерва, а также ветвей ganglion cervicale craniale. У собак, кроме того, идут тонкие ветви непосредственно из n. vagus или его rami pharyngei. Все названные ветви соединяются между собой, а у собак присоединяются к части plexus caroticus externus, при этом со значительными вариациями.
Glomus aorticum включает группы хромаффинных клеток, которые лежат на дуге аорты и нечетко отграничиваются от окружающих тканей. Эти клетки, как и клетки glomus caroticum регистрируют давление С02 в крови и передают информацию по ветвям n. vagus к ядрам продолговатого мозга. Значение и функции эфферентных волокон, которые заканчиваются в glomus caroticum и glomus aorticum, еще точно неизвестны.
Paraganglion aorticum abdominale располагается около вентральной поверхности aorta abdominalis и места возникновения a. mesenterica caudalis, и у новорожденных животных развит лучше, чем у взрослых. Его функции, также как и небольших групп хромаффинных клеток, например около п. tympanicus или у a. subclavia неизвестны.
Используемая литература: Анатомия собаки и кошки (Колл, авторов) / Пер. с нем. Е. Болдырева, И. Кравец. - М.: «АКВАРИУМ БУК», 2003. 580с., ил. цв. вкл.
Скачать реферат: