Циклические изменения волос
Волосяной покров собаки и кошки способен претерпевать изменения. Для его обновления и функционального приспособления к сезонной перемене погодных условий в волосяных фолликулах протекают эволюционные, трансформационные и инволюционные процессы. Эти процессы имеют циклический характер. Как следствие этого, необходимо знать различия между активными/анагенными и пассивиыми/телогениыми волосяными фолликулами. Трансформация анагенного волосяного фолликула в телогенный (инволюция волосяного фочликула) проявляется через возникновение временного катагенного фолликула. Получающийся из него в конце концов телогенный волосяной фолликул содержит телогенный (колбовидный) волос, который функционирует в составе волосяного покрова в течение нескольких месяцев (продолжительность зависит от вида и индивидуальных особенностей). К этому моменту в первичном волосяном фолликуле уже растет новый волос.
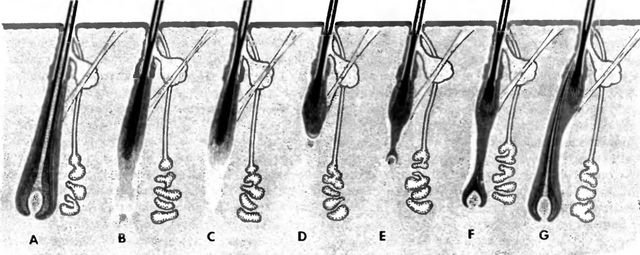
Рис. 8.
Циклические изменения волоса на примере первичного волосяного фолликула; A анагенный фолликул, В, С катаген-ный фолликул, D телогенный фолликул (с колбовидным волосом), Е — С телогенный фолликул с растущим зачатком волоса, "волосяным конусом" либо новым волосом
Красным цветом: внутреннее эпителиальное влагалище корня, синим цветом: наружное эпителиальное влагалище корня
Эти циклические процессы начинаются только после первой смены волос, которая у европейской лесной кошки происходит примерно через два месяца после рождения. Для домашней кошки до сих пор таких данных нет. Домашние собаки теряют детскую шерсть, в зависимости от породы, через три -шесть месяцев после рождения.
Активно растущий анагенный волосяной фолликул сравнительно глубоко проникает в кожу. Его наиболее характерным признаком является наличие волосяной луковицы. Последняя куполообразно охватывает соединительнотканный волосяной сосочек, обеспечивающий питание и иннервацию матрикса. В однослойном матриксе можно выделить — в определенной последовательности — несколько участков, ответственных за возникновение отдельных частей корня волоса и внутреннего эпителиального влагалища. Так, клетки мозгового слоя образуются центральными группами клеток матрикса, клетки коркового вещества и кутикулы корня — соответственно дальше расположенными клетками матрикса. Периферические клетки матрикса отвечают за формирование и рост всех трех слоев внутреннего эпителиального влагалища (кутикулы, внутреннего и наружного эпителиальных слоев). В наружном эпителиальном влагалище корня в этот активный период можно обнаружить значительное количество внутриклеточного гликогена, являющегося источником питательных веществ (Kligman, 1959; Zaun, 1968; Forslind, 1979; Schwarz, 1991).
После прекращения роста в анагенном волосяном фолликуле начинается фаза инволюции или катагенная фаза. Разрушение начинается с матрикса. Сначала прекращается образование клеток мозгового вещества. В результате этого корень волоса утоньшается Последовательно угасает активность и других клеток матрикса, которые все больше и больше отдаляются от волосяного сосочка. Таким образом, анагенный волосяной фолликул не просто выталкивается к поверхности кожи, как предполагалось раньше, а претерпевает серьезные инволюционные изменения, превращаясь в катагенный фолликул. Плотная мелкоячеистая капиллярная сеть в области луковицы также исчезает.
В результате разрушения анагенного волосяного фолликула в конце концов возникает телогенный волосяной фолликул. В этом фолликуле, уменьшившемся в зависимости от типа первичного волоса на половину или даже на две трети, распушенный в виде кисточки остаток корня волоса прочно закреплен в ороговевшем внутреннем эпителиальном влагалище. В результате этого возникает колбовидный волосяной фолликул (колбовидный волос), который со стороны дермы полностью окружен клетками редуцированного наружного эпителиального влагалища. Относительно того, сколько времени телогенный (колбовидный) волос сохраняется в коже, конкретных даных нет, но очевидно, что это зависит от типа волоса, вида животного и индивидуальных особенностей.
Сохраняющиеся вокруг телогенного волосяного фолликула остатки наружного эпителиального влагалища содержат клеточный материал, необходимый для образования нового волосяного фолликула. Это означает, что у взрослого животного образование нового анагенного волосяного фолликула происходит за счет эпителиальной/эпидермальной оболочки телогенного фолликула, а не за счет базального слоя эпидермиса, как у плода (Schwarz et al., 1986; Schwarz, 1991). Возникающая таким образом закладка волосяного фолликула, в которой уже можно распознать луковицу и сосочек, прорастает в виде эпителиального тяжа в дерму. Когда эпителиальный тяж достигает своей конечной, генетически обусловленной длины, матрикс волоса в нем уже бывает выделен и готов к формированию нового волоса. Если до этого пролиферация и рост были ориентированы в сторону подкожной основы, то теперь направление меняется на противоположное. Сформировавшие „конус" нового анагенного волоса клетки и внутреннее эпителиальное влагалище растут внутри эпителиального тяжа в сторону эпидермиса. Периферические клетки тяжа сохраняются и образуют наружное эпителиальное влагалище корня волоса. Телогенный/колбовидный волос выпадает только тогда, когда новый анагенный волос достигнет поверхности кожи или, как минимум, волосяного канала.
Смена волосяного покрова
На циклические изменения волоса влияют эндогенные и экзогенные факторы. Эндогенными факторами являются порода, пол, возраст и состояние здоровья, а также помимо всего прочего гормоны гипофиза, половых желез, коры надпочечников и щитовидной железы. К разряду внешних воздействий относятся свет, температура, питание, содержание и качество ухода.
Беременность и лактация вместе с гормональным изменениями могут привести у собаки и в некоторых случаях у кошки к диффузному выпадению телогенных волос через несколько недель после родов (Scott, 1990). Это происходит в результате того, что увеличивается и ускоряется переход анагенных волосяных фолликулов в телогенные, из которых затем волосы выпадают.
Решающее влияние на процесс смены волосяного покрова оказывает периодическое изменение длины светового дня. Животные, содержащиеся в естественных условиях, подвержены как циркадным, так и ритмическим годовым колебаниям. В основном складывается впечатление, что длина светового дня от 650 до 750 минут — как это бывает в Центральной Европе в марте и сентябре — является важнейшей предпосылкой для начала линьки. Линька у отдельных особей одной породы продолжается примерно одинаковое количество времени — от 43 до 53 дней, с небольшими отклонениями у отдельных пород собак.
Четко разграничить влияние света и температуры на процесс смены волосяного покрова невозможно. В настоящее время есть много данных, свидетельствующих о том, что изменения температуры в течение года играют роль лишь вспомогательного стимула и могут задержать начало линьки или увеличить ее продолжительноcть, например, весной (Торр, 1983).
В результате одомашнивания и селекции сезонная линька все больше уступает место постоянному обновлению волосяного покрова. Этот процесс продолжается в течение всего года с небольшим повышением интенсивности весной и осенью. Этому способствует, например, искусственное увеличение светового дня и почти постоянная температура при содержании животного в квартире. С другой стороны, обозначилась более выраженная тенденция к постоянной замене волос у некоторых пород собак, например, у боксеров, жесткошерстных и длинношерстных терьеров, шнауцеров, бриаров и спаниелей. Подобные наблюдения были также сделаны и в отношении кошек (Meyer et al., 1980; Торр, 1983; Schwarz/Neurand, 1986).
Однако по мере преобладания постоянной смены волосяного покрова над сезонной все больше и больше уменьшается значение взаимодействия между активными и пассивными волосяными фолликулами, необходимого для физиологического функционирования кожи. В результате этого вместо сезонной, но непродолжительной нагрузки кожа подвергается менее интенсивной, но практически постоянной. Это также снижает эффективность барьерной функции кожи и повышает риск кожных заболеваний (Schwarz, 1992).
САЛЬНЫЕ ЖЕЛЕЗЫ
Мешковидные, функционирующие по голокрин-ному типу сальные железы, glandulae sebaceae, встречаются на фолликулах первичных и вторичных волос во всех областях тела. Они возникают в период внутриутробного развития вместе с формированием корней волос, однако полностью завершают свое развитие только с наступлением половой зрелости. Размеры и форма сальных желез варьируют в зависимости от вида животного, породы, области тела и типа волосяного фолликула. Особенно крупные и дольчатые железы находятся на дорсальной стороне шеи, на спине и на хвосте, более мелкие и простые — на животе (Strickland/Calhoun, 1963; Meyer et al., 1981; Al-Bagdadi, 1993).
Все сальные железы, относящиеся к одной группе волос, расположены примерно на одном уровне, в сетчатом слое дермы. Периферические области желез состоят из мелких чашеобразных клеток с очень высокой митотической активностью. К центру железы клетки становятся крупнее и многообразнее, и в их цитоплазме накапливается все большее количество жировых капель. В конце концов клетки разрушаются и образуют in toto сальный секрет. Этот секрет в числе прочих веществ содержит стерилестер (различается в зависимости от вида животного), диэстер воска, свободные стеролы (холестерин) и фосфолипиды. Секрет сальных желез выводится в канал волоса, где смешивается с секретом апок-ринных трубчатых желез. На коже этот смешанный секрет образует липкую пленку, защищающую от ультрафиолетового облучения, поддерживающую эластичность кожи и волос и обладающую сильным водоотталкивающим действием, особенно у животных с густой шерстью. Деятельность сальных желез регулируется гормонально, особенно сильное действие оказывают андрогены. Изменения гормональных процессов, например, в связи с половым циклом, а также стрессы могут весьма существенно повлиять на секреторную активность желез. Функция сальных желез может быть нарушена бактериями и паразитами, проникающими в канал волоса и/или в сами железы. Пораженные участки кожи выглядят измененными и обычно очень болезненны (Nicolaides et al., 1968; Meyer/Neurand, 1976, 1977 с; Meyer et al., 1978 f; Sharaf et al., 1977; Schwarz et al., 1981).
АПОКРИННЫЕ ТРУБЧАТЫЕ ЖЕЛЕЗЫ
Помимо сальных желез в покрытой волосами коже собаки и кошки имеются еще апокринные трубчатые железы, glandulae tubuliformes apocrinae. Они существуют исключительно при фолликулах первичных волос. С этим связаны особенности распределения этих желез по телу. У кошки, например, в дорсальных областях кожи насчитывают от 1600 до 3200 желез/см2, а на животе — всего от 600 до 1300 желез/см2. У собаки плотность трубчатых желез наибольшая на шее и спине, а также на дистальных частях конечностей. На проксимальных частях конечностей, на голове и на боках плотность желез невелика. Меньше всего она на животе и внутренних поверхностях бедер. Плотность трубчатых желез и плотность пучков волосяных фолликулов не обязательно совпадают, так как не все пучки содержат первичные волосы; некоторые состоят только из вторичных волос и не содержат трубчатых желез (Claushen, 1933; Duchs, 1936; Brunsch, 1956; Jenkinson/Blackburn, 1968).
Апокринные трубчатые железы состоят из железистой ткани концевого отдела и выводного протока. Концевой отдел у собаки описывается как несильно извитая трубка или вытянутый клубок. У кошки она относительно небольшая и извитая. Размеры железистой части, особенно у собаки, различаются в зависимости от породы и области тела. У пород с длинной и тонкой шерстью они крупнее, чем у пород с короткой и грубой шерстью. Железистая часть располагается на уровне луковицы волоса, то есть на границе дермы и подкожной основы, в тупом угле, который волосяной фолликул образует с эпидермисом. Стенка концевого отдела образована слоем изопризматических и цилиндрических клеток, слоем миоэпителиоцитов, базальной мембраной и тонкой собственной пластинкой. Выводной проток апокринной трубчатой железы проходит почти параллельно волосяному фолликулу через мышцу, поднимающую волос, и открывается в волосяной канал над устьем сальной железы. Клетки выводного протока не поглощают ионы и не забирают, таким образом, воду из секрета (Clair, 1952; Lovell/Getty, 1957; Munger, 1965; Iwasaki, 1983; Meyer/Neurand, 1987).
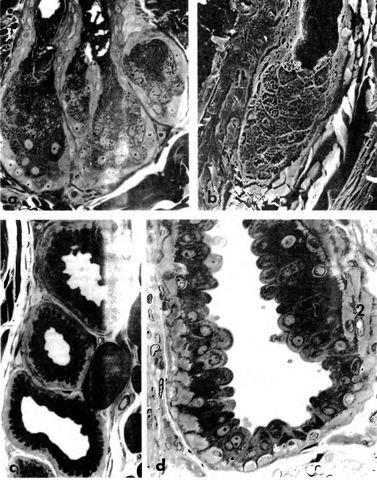
Рис. 9.
а, b Кошка — сальная железа (а 800-кратное увеличение, полутонкий срез, толуидиновый синий, b 880-кратное увеличение, REM); собака — с, d апокринная трубчатая железа, железистая часть, (с поперечный срез, 550-кратное увеличение, d продольный срез, 1350-кратное увеличение, полутонкий срез, толуидиновый синий)
1 железистый эпителий, 2 миоэпителиоциты
Апокринные трубчатые железы не иннервированы. Обмен веществ в них регулируется адренергическими субстанциями, что у пород с повышенной нервной возбудимостью может привести к усиленной и неспецифической секреции. В основе выделения секрета лежит релаксирующее действие, которое NO оказывает на миоэпителиоци-ты. Выделение этого вещества в секреторных клетках непосредственно зависит от интенсивности обмена веществ. При высоком уровне секреции большие количества приводят к релаксации мио-эпителиоцитов и, соответственно, к расширению просвета железы, и наоборот, меньшие количества NO вызывают прекращение выделения секрета, так как допускают сокращение миоэпителиоцитов с выдавливание секрета из полости железы. Вязкий, у собаки иногда водянистый, богатый гликопротеидами секрет на поверхности тела приобретает индивидуально специфический запах. Усиление секреции вследствие стресса часто приводит к появлению слишком сильного запаха, который при влажной шерсти может ощущаться даже человеком. Апокринные трубчатые железы ни филогенетически, ни онтогенетически, ни с точки зрения их морфологии и функции не соответствуют эккринным потовым железам высших приматов. В терморегуляции они не играют никакой роли. Поэтому совершенно ошибочно называть их потовыми железами, glandulae sudoriferae (Maezulli/Callahan, 1957; Cotton et al., 1975; Neurand et al., 1977 b; Meyer et al., 1978 c, 1987, 1991, 1994; Meyer/Schwarz, 1992).