Каждый эколог, как и биолог любой другой специальности, должен определить свое отношение к понятию «вид», с которым связаны все другие понятия и проблемы данной науки. В последние годы на страницах советских биологических журналов оживленно дискутируется вопрос о виде и видообразовании, всколыхнувший широкие круги ученых. Данная проблема слишком обширна, чтобы мы могли сколько-нибудь подробно осветить ее в рамках нашей книги, однако считаем необходимым сделать краткие замечания по поводу воззрений Ч. Дарвина и Т. Д. Лысенко на этот счет, которые в настоящее время нередко противопоставляются в качестве двух взаимоисключающих концепций.
«В учении Дарвина я согласен с теорией развития, дарвиновский же способ доказательства (борьба за существование, естественный отбор) считаю всего лишь первым, временным, несовершенным выражением только что открытого факта» — заявил Ф. Энгельс 12 сентября 1875 г. П. Лаврову, и уже вскоре после этого сам Дарвин в письмах к М. Вагнеру (1876), - М. Неймайру (1877), К. Земперу (1881) и другим естествоиспытателям писал о своей «величайшей ошибке», выразившейся в недоучете прямого действия условий жизни на организм, независимо от естественного отбора.
Теория видообразования Дарвина построена на признании ряда устаревших и односторонних положений, которые теперь или нельзя принять совершенно, или являются спорными и недоказанными. К ним относятся: неопределенный, случайный характер изменчивости, постепенность перехода индивидуальных отличий в видовые, приводящая к представлению о нереальности вида, и отрицание скачков в природе; связывание видообразования с перенаселенностью (мальтузианство, по Энгельсу), которая якобы неизбежно вытекает из геометрической прогрессии размножения; игнорирование прямого влияния условий жизни на организм—«... без наличия такого перенаселения: например, при переселении растений и животных в новые места, где новые климатические, почвенные и прочие условия вызывают изменение» (Ф. Энгельс, Диалектика природы, 1946, стр. 250). Вследствие этих существенных недостатков теория Дарвина, как она была сформулирована в 1859 г., теперь полностью принята быть не может, что ясно видел и ее творец уже в 70-х годах прошлого столетия.
Работами биологов-материалистов теория видообразования значительно усовершенствована и современный творческий дарвинизм позволяет дать наиболее правильную и всеобъемлющую концепцию вида. Вместе с тем необходимо не смешивать позицию в этом вопросе творческого дарвинизма с некоторыми субъективными взглядами Т. Д. Лысенко, высказанными им в 1950 г.
Так, спорными в представлениях Т. Д. Лысенко мы считаем отрицание им дивергенции, игнорирование ареала в определении вида, непризнание наличия переходных форм между видами в ископаемом состоянии, допущение возможности возникновения существующих видов сызнова и их происхождения из разных источников, наконец, отнесение к проблеме видообразования случаев разового перерождения некоторых культурных растений в сорные, которые могут объясняться другими причинами. Однако все эти моменты не имеют столь большого значения, какое им приписывается некоторыми биологами, призывающими полностью отказаться от достижений творческого дарвинизма и вернуться к теории Дарвина в ее первоначальном виде.
Считая эти положения сомнительными, мы в то же время признаем во взглядах Т. Д. Лысенко о виде и видообразовании наличие ряда ценных и плодотворных соображений относительно объективной реальности видов, их качественной обособленности, о разновидностях как формах существования вида, о специфике межвидовых и внутривидовых отношений организмов, о изменении условий жизни как исходной причине видообразования. Однако, к сожалению, некоторые биологи еще не оценили в полной мере значения перечисленных моментов, которые представляют не только элементы теоретической схемы, но и инструмент, которым можно удобно пользоваться в систематике, экологии, биогеографии и других биологических науках.
Примеры перерождения культурных растений и порождения ими специфических сорняков справедливо вызывают критическое отношение к себе со стороны ряда биологов, как не связанные прямо с проблемой видообразования. Вместо образования нового и поступательного движения вперед, каким только и может быть процесс видообразования, здесь имеет место «порождение сызнова» существующего, т. е. определенное движение по кругу. Вызывает сомнение также то, что одни и те же виды порождаются различными исходными, что открывает двери принципу полифилии, который враждебен дарвинизму. Вряд ли широко осуществляется в природе положение, согласно которому родительские виды сразу рождают новые дочерние, иногда относящиеся даже к другим родам.
Исходя из основного закона биологии, следует говорить о соответствии видовой формы условиям жизни. Мы полагаем, что в соответствии с характером изменения условий жизни (но не среды вообще) и видовая форма может измениться, превратиться в другую внезапно или постепенно. Имеющиеся во втором случае переходные формы исчезнут без всякой внутривидовой конкуренции 61 вследствие изменения условий жизни, которым они больше не отвечают.
Следует заметить, что спорность вопроса, можно ли теорию видообразования творческого дарвинизма обосновать малодоказательными в этом отношении фактами перерождения культурных растений, привела в 1952 г. некоторых биологов (Н. В. Турбин, Н. Д. Иванов) к попытке ревизовать самые основы учения И. В. Мичурина. Спорными вновь объявлены вопросы о том, были ли у Дарвина ошибки или он диалектико-материалистически трактовал эволюцию, считал ли он вид реальным или нереальным, вредно ли мальтузианство в биологии, адэкватна ли изменчивость наследственности внешним воздействиям и: т. п.
Совершенно неправильно, как делают некоторые, рассматривать порознь эволюционное и революционное развитие, количественные и качественные изменения. Теория Дарвина представляет собой именно эволюционную концепцию, в отличие от творческого дарвинизма, составляющего теорию развиты я органического мира.
По основному вопросу «происхождение видов» позиция творческого дарвинизма представляет диалектический синтез прежних теорий. «Реальность и неизменность» видов у Линнея, «нереальность и изменяемость» видов у Ламарка и Дарвина синтезируются в современном представлении как «реальность и изменяемость» видов. Но и «реальность» и «изменяемость» видов в диалектико-материалистической биологии совсем не те, какими эти понятия были в науке прошлого. «Реальность» вида по Линнею — креационистская, результат творческого акта; теперь — материалистическая, признание вида объективно существующим явлением природы. «Изменяемость» вида по Ламарку, Дарвину — метафизическая, медленный процесс постепенных количественных изменений; теперь — диалектико-материалистическая, основанная на признании внутренней связи количественных и качественных изменений.
Теория эволюции Дарвина господствует в биологической науке почти сто лет. За этот период не было недостатка в попытках замены ее автогенезом или эктогенезом, ортогенезом, номогенезом или гетерогенезом и т. п., но все названные и многие другие «теории» эволюции, построенные на отрицании естественного отбора, неизбежно терпели крах как идеалистические и метафизические. Развиваемая ныне диалектико-материалистическая теория развития органического мира преодолевает ошибки и ограниченность учения Дарвина и сохраняет все ценное из дарвинизма. Поэтому несомненно, что творческий дарвинизм представляет новую, более высокую ступень в поступательном движении биологической науки.
С позиций творческого дарвинизма возможно и необходимо разработать учение о виде.
В истории биологии попыток формулировать понятие «вид» было очень много, по существу столько, сколько биологов им занималось. В. Л. Комаров совершенно верно сказал: «Если просто расположить определения вида, данные ему различными учеными в различное время, в хронологическом порядке, то из этого ничего полезного не выйдет. Получится нагромождение мало понятных фраз, из которых читатель ничего не вынесет. Учеными в их работе руководили как состояние науки, так и социальная среда. Различен был материал, над которым они работали, различны и мысли, которые их питали. Различна также была и цель, которую они себе ставили в своих исканиях».
Вид представляет собой сложное биологическое явление, и потому стремление отразить в одной формулировке вое его стороны приводит к усложнению и громоздкости определения.
К недостаткам известной формулировки вида, данной Т. Д. Лысенко, относится то, что он не подчеркивает в числе признаков вида наличие определенного ареала, главный же упор делает на специфике внутривидовых отношений, характеристика которых исследователю в большинстве случаев недоступна.
Мы полагаем, что правильное представление о виде может дать следующее наше определение.
Вид —реально существующая в природе совокупность организмов характерного строения и внешнего облика, исторически развившаяся от общих предков; соответствующая условиям жизни в пределах свойственного ей ареала и поддерживаемая в своих признаках определенным типом обмена веществ; дающая в ряде поколений сходное потомство, в котором, в меру разнообразия условий жизни, одновременно проявляется консерватизм наследственности и внутривидовая изменчивость; качественно обособленная от других видов отсутствием переходов среди живых представителей, при их возможном сохранении в ископаемом состоянии; превращающаяся в другой вид, в зависимости от характера изменения условий жизни, медленно и постепенно или быстро и резко.
Со времен Линнея вид рассматривается не как нечто неделимое, состоящее из вполне тождественных особей, а как группа особей, в известной мере между собою различающихся. В связи с этим встает вопрос о внутривидовых категориях, которые иногда неправильно называют «таксономическими единицами», неправильно, так как действительной таксономической единицей является только вид.
Следует сказать, что как последователи креационизма, так и сторонники эволюционизма в равной мере допускают существование внутри вида разновидностей. Однако понимают вид, разновидность и соотношение между ними те и другие совершенно по-разному.
Для креационистов вид постоянен и изменяться не может. В то же время под влиянием внешних условий особи вида в какой-то мере могут между собой различаться. Изменения особей в общем незначительны, временны и преходящи, они прекращаются вместе с исчезновением породивших их условий. С этой точки зрения виды, реально существующие в природе, качественно различны, разновидности же отличаются от вида лишь количественно.
Таксономически такое соотношение вида и разновидности выражается так, что вид обозначается по латыни биноминально, а разновидность — триноминально. Например, моллюск ушковый прудовик Radix auricularia (L. ) имеет ряд разновидностей, в том числе var. ampla Наrtm., var. fluviatilis Sсhad. и др. Выходит, что вид существует как бы вне своих разновидностей, а последние — независимо от него.
Когда в биологических науках победил эволюционный взгляд, казалось, что вопрос о виде и разновидности должен разрешиться как-то по-иному. Но Дарвин не сумел найти объяснения эволюционному процессу, который одновременно характеризуется изменением видов, превращением одних видов в другие, и в то же время наличием разрывов между видами. Разновидности, по Дарвину, и выполняют эту весьма сложную роль: в качестве «ступенек» они содействуют превращению одного вида в другой, а затем поспешно исчезают в результате «внутривидовой конкуренции».
Таксономическое соотношение вида и разновидности и в этом случае оставалось таким же, как при Линнее: разновидность, составляя часть вида, по существу оказывалась за его пределами. По Дарвину, вид существует и процветает, как бы не имея разновидностей. Но под влиянием «перенаселения и внутривидовой конкуренции» происходит нарастание различий, именуемое дивергенцией, и вид распадается — происходит образование двух или нескольких новых видов. Следовательно, появление разновидности означает, по Дарвину, начало распада вида, разновидность находится как бы за рамками вида, на пути между двумя видами. По Дарвину, даже можно проследить степень «продвижения» разновидности на ее пути от старого вида к новому, проявляющуюся в нарастании морфологических отличий.
Ч. Дарвин писал: «... я считаю индивидуальные различия, хотя они мало интересны для систематика, крайне важными для нас, так как они представляют собой первые шаги к образованию разновидностей, настолько незначительных, что о них, как обыкновенно полагают, не стоит даже упоминать в естественно-исторических сочинениях. Разновидности, которые в некоторой степени более различаются между собой и в некоторой степени постоянны, я рассматриваю как ступени к более резко выраженным и постоянным разновидностям, а эти последние—как ступени к подвидам, а затем к видам. Переход с одной ступени различия на другую во многих случаях мог представлять собой простой результат особенностей самого организма и различных физических условий, которым он долго подвергался но по отношению к важнейшим приспособительным признакам переход с одной ступени на другую можно с уверенностью приписать накопляющему действию естественного отбора, ...а равно действию увеличивающегося упражнения или неупражнения органов. Ясно выраженная разновидность может быть вследствие этого названа зарождающимся видом...
Из всего сказанного ясно, что термин вид я считаю совершенно произвольным, придуманным ради удобства, для обозначения групп особей, близко между собою схожих, и существенно не отличающимся от термина разновидность, которым обозначают формы, менее резко различающиеся и более колеблющиеся (в своих признаках). Также и термин разновидность, в сравнении с просто индивидуальными различиями, применяется произвольно, ради удобства».
Такова была точка зрения Дарвина. Условность различий между видом и разновидностью, разницу между которыми он считал чисто количественной, а переход от одного вида к другому — исключительно постепенным, приводила к порочному представлению о нереальности вида как явления природы.
В результате этого, если следовать взглядам Дарвина, живая природа должна представлять собой весьма странную картину. В ней постоянно имеются, наряду с ограниченным числом определенных видов («хорошие виды» систематиков), преобладающие количественно формы, видовая принадлежность которых точно установлена быть не может: разновидности разных степеней различия занимающие положение между различными видами («сомнительные виды» систематиков).
Ошибки Дарвина в трактовке вопросов видообразования усугублялись систематиками, которые в практической работе не могли руководствоваться «относительностью» различий между видом и разновидностью, о которой писали дарвинисты, и вынуждены были искать каких-то более определенных критериев для установления этих различий.
Изменчивость организмов всегда представляла и представляет много хлопот систематикам. Отсюда стремление некоторых как-то сузить понятие вида, избежав тем самым «излишней» вариабильности особей.
Своеобразный «выход» из этого положения нашел лионский купец А. Жордан, интересовавшийся ботаникой. «По Жордану, следует признавать за самостоятельный вид всякое растение, обладающее признаком, передающимся по наследству, как бы ничтожен не был этот признак». Как известно, Жордан исследовал невзрачное растение — крупку весеннюю Draba verna L. После десятилетних наблюдений он разбил этот линнеевский вид на 10 видов; через двадцать лет он различал их 32, а через тридцать лет — уже целых 200.
Подобное течение в систематике, получившее название жорданизма, или «распыливания видов», основывается на ложной попытке найти предел изменчивости и считать видом такую группу особей, которая характеризуется постоянством признаков. Взгляды Жордана разделял Де-Фриз, который также полагал, что каждая мутация энотеры представляет новый вид.
Жорданисты чрезвычайно осложняют систематику организмов, так как вид сводят не только до разновидности, но и индивидуальной изменчивости. Всякое, самое незначительное отличие, являющееся наследственным, считается ими достаточным для установления нового вида.
От прежних жорданистов прямые нити тянутся к современным систематикам-ботаникам. Ряд биологов обращает внимание на неправильное понимание многими ботаниками понятия вид. По взглядам ботаников, «вид представляет собою множество подобных один другому организмов, свойства которых наследственны и передаются из поколения в поколение».
Как указывает Л. С. Берг, на основании этого определения любую наследственную форму, как бы она ни была близка к соседней, можно описывать как особый вид. Так ботаники и делают, беспрерывно описывая все новые и новые виды. Этому процессу — бесконечному увеличению ботанической номенклатуры, никогда не будет конца, ибо следует твердо помнить, что в каждом новом местонахождении обитает своя, особая форма растения (и животного). Если пока данное положение не доказано для всех видов, это есть следствие нашей плохой способности к различению. Но так как познавательные способности человека с течением времени совершенствуются, то постепенно умение различать особенности возрастает.
С этим прямо соглашается крупнейший представитель ботанической систематики В. Л. Комаров: «Линнеон то же, что вид, но вначале он был крупнее, теперь же, с усвоением более тонкой диагностики, стал много уже». Из этого и других замечаний В. Л. Комарова следует, что в практической деятельности систематика он нередко подходил к понятию вид субъективистски.
Неправильное понимание частью современных ботаников-систематиков понятия «вид» приводит к тому, что они не принимают диалектико-материалистической теории видообразования.
По-иному подходит к вопросу большинство зоологов, которые сумели быстро разоблачить попытки антиисторической трактовки вида некоторыми учеными и стоят на принципиально правильном пути.
Известный русский зоолог А. П. Семенов-Тян-Шанский в работе «Таксономические границы вида и его подразделений» (1910) впервые подробно характеризовал внутривидовые категории. Таковые он подразделил на две группы: географические (подвид, или раса, и племя) и негеографические (морфа и аберрация). Термин «разновидность», которым слишком злоупотребляли и в зоологии, и в ботанике, при этом совсем отпал. Им стали пользоваться в тех случаях, когда нельзя точно определить таксономическое значение внутривидовой формы. Крупный знаток систематики Л. С. Берг (1948) развил идеи А. П. Семенова-Тян-Шанского применительно к рыбам следующим образом.
Видом (species), или линнеоном, называется совокупность особей, занимающих определенную географическую область и обладающих рядом определенных признаков, передаваемых по наследству и всегда отличающих данный вид от близких видов. Подвидом (subspecies) называется совокупность особей, связанная с близким к ней видом или расой наличностью переходных форм, но имеющая определенную географическую область обитания. Племя (natio) представляет как бы подвид второго порядка.
В отличие от рассмотренных географических понятий, негеографические подразделения вида (экотип, биотип, морфа, аберрация) не связаны с какой-либо определенной областью обитания, а возникают под влиянием тех или иных внешних условий. Экотип (infraspecies), или раса, элементарный вид, жорданон представляет стойкую форму, связанную с видом переходными признаками, но отличимую по комбинации признаков (расы домашних животных и культурных растений). Биотип — более мелкая категория. Морфа (morpha) —уклонения от основной формы, вызванные изменением внешних условий. Аберрация — индивидуальные наследственные изменения (меланизм и пр. ).
Существенная ошибка системы Л. С. Берга заключается в том, что в ней вид не существует в форме разновидностей, но сам представляет, наряду с подвидом и племенем, лишь одну из географических таксономических единиц. На практике при классификации пресноводных рыб СССР это вело к тому, что Л. С. Берг называл плотвой Rutilus rutilus (L. ) не весь этот вид, обнимающий 8 подвидов, а лишь европейский подвид, ельцом Leuciscus leuciscus (L. ) не весь вид, включающий европейский и сибирский подвиды, а лишь первый и т. д.
Однако основное и плодотворное в классификации А. П. Семенова-Тян-Шанского и Л. С. Берга состоит в том, что вид и «разновидности» (т. е. различные географические и негеографические подразделения вида) — это не одно и то же. Усовершенствование этой классификации заключается в том, чтобы выделить вид из числа географических единиц и считать, что именно вид состоит из географических и негеографических форм. Можно и нужно спорить по вопросу о характере внутривидовых подразделений, но неправильно поступать так, как это делают многие ботаники: вид смешивают с подвидом, а в остальном не разбираются, называя это разновидностями.
Совершенно очевидно, что вилы животных, растений и микроорганизмов не равновелики и не равноценны. В каждой области частной систематики о этом отношении будут свой подход и свои критерии. А. П. Семенов-Тян-Шанский (1910) совершенно правильно писал, что «... вполне эквивалентными виды никогда быть не могут. Будучи вершинными в данный момент побегами или же концами ветвей сложного дерева, виды в каждой группе могут быть весьма между собою различны и по своему геологическому возрасту, и по происхождению, т. е. по степени родства с наиболее близкими к ним из ныне живущих форм и по темпу, в котором они обособлялись и слагались, и по характеру факторов, которым они обязаны своим происхождением, и по пути, который они прошли, что-бы придти к современной форме».
Неравноценность видов разных групп организмов и разная степень их изученности будет иметь своим следствием разную детальность разработки системы внутривидовых подразделений, но принципы выделения вида (в смысле объема этого понятия) должны быть общими. Сейчас этого еще нет.
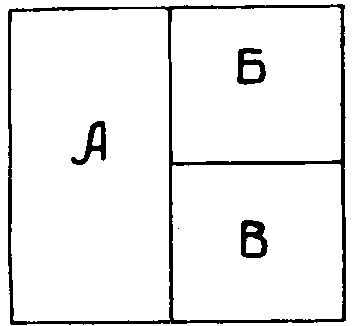
Рис. 1. Схема, иллюстрирующая прежнее и современное представление о соотношении вида и разновидностей (в смысле таксономическом и ареалогическом). Прежнее представление: А—вид, Б и В — подвиды. Современное представление: А, Б, В — формы существования вида.
Неправильное применение в орнитологии географического критерия привело к представлению о круге форм. Клейншмидт заменил, линнеевский вид понятием круга форм (Formenkreis). Признавая обычную ограниченную эволюцию в пределах каждого круга форм, этот автор в 1930 г. защищал представление о возникновении каждого круга форм путем особого творческого акта.
В. Л. Комаров, стремясь избежать отмеченных выше трудностей систематической работы, создал учение о рядах, которое положено в основу грандиозной «Флоры России». Согласно этому учению подвиды растений нередко описываются в качестве самостоятельных видов, а настоящий вид именуется «рядом», который по существу недалек от «круга» Клейншмидта.
В. Л. Комаров обосновывает свое учение о рядах следующим образом. «Там, где ранее признавался один вид, новое более тщательное исследование нередко обнаруживает несколько близких видов, занявших каждый определенное место в экономике природы. Все эти виды получают самостоятельные названия, но перед всей их группой следует ставить слово series — ряд». «Важно не то, признавать ли данную форму за разновидность, подвид или вид, а то, чтобы показать родство близких форм между собой».
Таким образом, В. Л. Комаров исходит из неверной дарвинской формулы об относительности различий между видом и разновидностью, причем без всяких к тому оснований видит в «учении о рядах» какое-то усовершенствование систематики. Совсем напротив!
Л. С. Берг (1950), разбирая недостатки ботанической номенклатуры и понятия вида у ботаников, вполне резонно подчеркивает значение гиатуса. Между двумя видами обязательно должен существовать диагностический перерыв. Поэтому определение вида недостаточно строить лишь на подчеркивании наследственности признаков, но нужно также указать ряд определенных признаков, всегда отличающих данный вид от близких видов. Если это условие не осуществляется, если по диагностическим признакам одна географическая наследственная форма постепенно переходит в другую, то мы имеем дело не с двумя видами, а с двумя подвидами одного вида. Как указывалось выше, при определении сущности явления имеет значение не столько связь его с другими явлениями, сколько отличие от них.
Понимание вида в «узких границах» неминуемо ведет не только к излишнему дроблению и увеличению числа систематических единиц, но и субъективизму. Постоянное нахождение новых отличии и переходных форм делает работу систематика, стоящего на таких позициях, совершенно бесперспективной, так как изучить видовой состав какой-либо группы организмов по существу становится невозможным. Напротив, систематик, исходящий из «широкого» понимания вида, остается на строгой почве фактов: границы между видами оказываются резкими, качественными, а многочисленные формы, часто связанные переходами, рассматриваются как разновидности и для последних достаточно описания лишь наиболее отличных. В этом случае работа систематика приобретает необходимые в науке объективные критерии и опирается на факты, причем сходные выводы могут быть сделаны разными исследователями, что совершенно невозможно в первом случае. Описание видов -— исторически выполнимая задача; описание же бесконечного количества разновидностей следует заменить изучением закономерностей их образования.
Стремление к измельчению видов свойственно не только ботаникам, но и части зоологов. Особенно усердствуют в этом отношении американские зоологи. Бурый медведь (Ursus arctos L. ), распространенный в Европе, Северной Африке, Северной Азии и в Северной Америке, образует множество местных форм. Н. А. Бобринский, Б. А. Кузнецов и А. П. Кузякиш (1944) отмечают в составе фауны России шесть подвидов бурого медведя: обыкновенного (Ur. аr. arctos L. ), берингийского (Ur. ar. beringianus Midd. ), манчжурского (Ur. ar. mandchuricus He tide), кавказского (Ur. ar. caucasicusi Smir. ), сирийского (Ur. ar. syriacus H. et. Ehr. ) и белокоготного (Ur. ar. leuconyx Sev. ).
В то же время из Северной Америки описано 95 форм бурого медведя, в том числе целый ряд особых видов. У ботаников в аналогичном случае получилось бы более 100 отдельных видов.
Взгляды Т. Д. Лысенко о соотношении вида и разновидностей дают систематикам возможность дальше плодотворно разрабатывать данную трудную область науки. Разновидности — это формы существования данного вида, а не ступеньки его превращения в другой вид. Появление разновидности — не начало конца вида, как следовало по Дарвину, но, наоборот, его упрочение. Чем больше разновидностей имеет тот или другой вид, чем разнообразнее внутривидовые популяции, тем больше обеспечиваются, хотя бы, например, перекрестным оплодотворением, возможности процветания вида и всех его разновидностей.
В этой постановке вопроса разновидности номенклатурно должны стоять не «вне» вида, а «внутри» его. Если возьмем пример с прудовиками, то мы должны сказать, что вид Radix auricularia представлен следующими формами: Radix auricularia auricularia (но не typ. ), R. a. ampla и т. д.
Учитывая необходимость характеризовать вид через формы его существования, следует заменить бинарную номенклатуру тринарной, как это уже и сделано в ряде отраслей зоологии.
Вопрос о характере внутривидовых подразделений приобретает с позиций творческого дарвинизма не просто номенклатурный, но особый биологический интерес. С этой точки зрения необходимо изучение разных групп организмов. Сейчас можно с уверенностью сказать о необходимости различать среди разновидностей по крайней мере подвиды ( географические формы) и морфы (негеографические формы).
Понятие вид, как и проблема видообразования, — предмет не только систематики, но биологии в делом. В подтверждение этого укажем основные биологические аспекты вида: систематический — реально существующая в природе совокупность, сходных особей, в зависимости от условий жизни представленная одной, несколькими или многими формами существования; морфологический — сходство строения и внешнего облика особей вида сочетается с наличием значительной изменчивости — индивидуальной и групповой (половой, возрастной, размерной, сезонной, биотопной, географической), при отсутствии в живом состоянии промежуточных особей между разными видами (наличие разрыва между ними — гиатуса) физиологический — свободное скрещивание особей одного вида и его затруднение между разными видами, с образованием в большинстве случаев бесплодных гибридов; экологический—определенный уровень численности особей, сложная динамика численности организмов в связи с занятием особями вида определенного местообитания, соответствующего необходимым условиям жизни, при наличии качественного отличия внутривидовых отношений от межвидовых; генетический—консерватизм наследственности и ее изменчивость, в зависимости от стабильности и динамики условий жизни; биогеографический — наличие у вида определенного ареала (сплошного или разорванного), исторически изменяющегося под влиянием перемен в условиях жизни и соответствующего изменения численности популяции; возможность существования в одной местности близких видов и нескольких негеографических форм («экологических», «биологических») одного вида, при наличии географического замещения (викариата) у подвидов; палеонтологический — наличие предков в ископаемом состоянии и возможность связи с ними посредством промежуточных форм; исторический — в индивидуальном развитии особи отражается история развития предков и закладываются пути развития потомков.
Трудности практического использования всех возможных критериев определения вида, с проведением всеобъемлющего действительно биологического анализа, ведут к тому, что систематики зоологи и ботаники часто ограничиваются изучением лишь двух сторон —морфологии и распространения.
Экологи внесут большой вклад в дальнейшую разработку учения о виде, если глубоко и всесторонне изучат экологический аспект проблемы, рассматривая материал под углом зрения наличия в природе разных путей видообразования.
Используемая литература: Основы Экологии: Учеб. лит-ра./Б. Г. Иоганзен
Под. ред.: А. В. Коваленок,-
Т.: Типография № 1,-58 г.
Скачать реферат:
Пароль на архив: privetstudent.com