КОНЕЧНЫЙ мозг
Конечный мозг, telencephalon, у млекопитающих формирует большую часть мозга, который настолько доминирует морфологически, что покрывает до мозжечка все остальные участки головного мозга, так как даже функционально в иерархической организации центральной нервной системы стоит на первом месте. Поэтому эта часть головного мозга называется большим мозгом, cerebrum. Конечный мозг парный и состоит из двух полушарий, которые лежат над стволом мозга с плащом, pallium, и над базальной частью, ганглиевым холмом (с полосатым телом, corpus striatum) соединяется с промежуточным мозгом, а через него — со всеми другими частями мозга.
В филогенезе различают три фазы развития частей конечного мозга: paleopallium — древнейший участок, воплощает вместе с более молодым участком, archipallium, обонятельный мозг; между обоими проходит neopallium, который претерпевает значительное изменение и более древние отделы мозга оттесняет базально и медиально.
Обонятельный мозг, rhinencephalon, отделяется от neopallium латеральной обонятельной бороздой, sulcus rhinalis lateralis. Он начинается рострально выделяющейся, особенно у собак, обонятельной луковицей, bulbus olfactorius. Её поверхность покрыта обонятельными нитями, fila olfactoria, которые проходят через продырявленную пластинку, lamina cribrosa, решётчатой кости и идут в обонятельные клетки слизистой оболочки носа (обонятельные рецепторы).
Каудально к луковице примыкает обонятельный тракт, tractus olfactorii, который у собак проходит в виде двух белых тяжей (tractus olfactorius lateralis et medialis), а у кошек поверхностно отмечается только tractus olfactorius lateralis. Обонятельный тракт покрыт серым веществом, которое латерально продолжается в виде латеральной обонятельной извилины, gyrus olfactorius lateralis, а медиально — в виде медиальной обонятельной извилины, gyrus olfactorius medialis. Обе извилины ограничивают треугольное поле, обонятельный треугольник, trigonum olfactoriurn, которое у плотоядных животных вентрально выдаётся в виде обонятельного бугорка, tuberculum olfactoriurn. Его каудальная часть формирует идущую с медиальной поверхности полушарий диагональную извилину, gyrus diagonalis.
Gyrus olfactorius lateralis идёт каудально в грушевидную долю, lobus piriformis. Стволовая часть обонятельного мозга — миндалевидное ядро, corpus amygdaloideum, относится к ганглиевому холму. Ядро соединяется с гиппокампом, а также посредством концевой полоски, stria terminalis с area septalis.
Связь половин обонятельного мозга осуществляется через ростральную спайку, commissura rostralis.
Грушевидная доля, lobus piriformis, является важной областью восприятия в обонятельном мозге. corpus amygdaloideum относится к лимбической системе.
Archipallium в процессе филогенеза оттесняется медиальной стенкой полушарий и характерно свёртывается при расширении neopallium. Одновременно эта область дугообразно вытягивается каудо-вентрально и из-за своей формы называется гиппокампом, hippocampus, или аммоновым рогом, cornu ammonis.
У домашних млекопитающих поверхность гиппокампа гладкая, лишена характерных структур, которые присутствуют у человека. У плотоядных животных особенно отмечается форма гиппокампа в виде бараньего рога, что обусловливает его название.
Поверхностно лежащая окологиппокамповая извилина, gyrus parahippocampus, сворачивается у борозды гиппокампа, sulcus hippocampi, причём на конце маленького завитка располагается зубчатая извилина, gyrus dentatus. Кора лежит под мозговым листком, лотком гиппокампа, alveus hyppocampi (-/4), который медиально переходит в бахромку, fimbria hippocampi (-/5). Fimbriae собираются в ножки свода, crura fomicis, которые сначала идут раздельно, далее рострально выдаются в виде тела свода, corpus fomicis в III желудочек, погружаются в виде столбов свода, columnae fomicis, в латеральную стенку III желудочка и заканчиваются в сосцевидном теле, corpus mamillare.
Формирование и увеличение объёма спайки neopallium, мозолистого тела, corpus callosum, отделяет гиппокамп от рострального участка archi pallium, паратерминальной извилины, gyrusparaterminalis, которая с примыкающим подмозолистым полем, area subcallosa относится к прекомиссуральному полю, area praecommissuralis. Она одновременно формирует тонкую медиальную стенку бокового желудочка. Мозолистое тело покрыто тонким слоем коры archi pallium, indusium griseum.
Свод, fornix, граничит вентрально с мозолистым телом, но отделяется от него на переходе в дугообразные columnae fornicis тонкой перегородкой (area postcommissuralis, прозрачной перегородкой, septum pellucidum), объём которой зависит от величины мозолистого тела с ростральной стороны. Septum pellucidum чаще всего лежит по средней линии. У плотоядных животных рост перегородки происходит неполностью, вследствие чего образуется полость, которая у кошек с ростральной стороны может быть открыта (Thompson, 1932). Так называемая полость прозрачной перегородки, cavum septi pellucidi, не имеет ничего общего с желудочком мозга.
В зависимости от строения neopallium большой мозг собак и кошек имеет извилины, а, например, у грызунов и насекомоядных гладкий. Головной мозг всех млекопитающих имеет следующие борозды: продольная щель большого мозга, fissura longitudinalis cerebri (между полушариями), sulcus hippocampi, латеральная обонятельная борозда, sulcus rhinalis lateralis (между palaeopalliumn neopallium), а также sulcus endorhinalis (между tuberculum olfactorium и tractus olfactorius lateralis). Таким образом, извилистое строение головного мозга обусловлено строением neopallium.
Количество извилин головного мозга обусловливается не только ступенью развития, но и размером животных, который обусловливает увеличение cortex cerebri. У крупных животных, стоящих на высокой ступени развития (выражена психическая иерархия, работоспособность головного мозга), neopallium имеет большее количество извилин.
На извилистом головном мозге всегда присутствует латеральная сильвиева борозда, fissura sylvia (lateralis cerebri)
Она располагается в зоне роста, от которой дугообразно рострально и каудально растёт neopallium и укладывается в складки, в то время как зона роста сохраняется в виде островка (insula, островок Рейля) и погружена в глубину.
В отличие от копытных животных, у плотоядных островок не собирается в складки и не покрыт бороздами. Потому эта борозда у собак и кошек называется fissura pseudosylvia. В данной книге авторы придерживаются названия fissura lateralis cerebri, или fissura sylvia. Различием между простым (у плотоядных) и имеющим большое количество извилин головным мозгом в том, что в последнем случае погружение коры островка происходит с формированием вентролатерально лежащей ямки, fossa lateralis (sylvii), в то время как в головном мозге плотоядных вся кора лежит поверхностно и разделяет фронтальную и височную доли поверхностной бороздой, fissura lateralis. Остальные топографические названия (зона роста neopallium, ограда, ганглиевый холм) сходны.
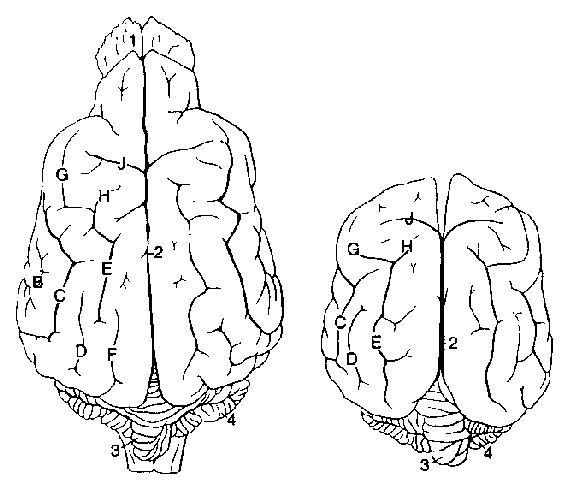
Рис. 11 Внешний вид головного мозга: слева представлен головной мозг длинноголовой собаки (немецкой овчарки), а справа — короткоголовой (французский бульдог) (по Seiferle, 1992)
В sulcus ectosylvius; С sulcus suprasylvius; D sulcus ectomarginalis; E sulcus marginalis; F sulcus endomarginalis; G sulcus coronalis; H sulcus ansatus; J sulcus cruciatus
1 bulbus olfactorius; 2 fissura longitudinalis; 3 vermis cerebelli; 4 hemisphaerium cerebelli
Островок также является областью коры, которая обеспечивает соединение neopallium с промежуточным мозгом. Эта область развивается в многоклеточный участок, который называется ядра, nuclei, или ганглии конечного мозга. Вместе с непосредственно присоединяющимся ядром промежуточного мозга (globus pallidum) эти области формируют так называемые базальные ганглии. К ним также относится corpus amygdaloideum, ядерная область palaeoencephalon.
Полушария большого мозга собак и кошек выделяются относительно простой схемой извилин и борозд. Преобладают дугообразные и вертикальные извилины, боковые выражены незначительно.
У собак после рождения происходит удлинение головного мозга, особенно лобных долей, и перемещение bulbi olfactorii рострально. Кроме того, увеличиваются борозды. Sulci у новорожденных только обозначены. Имеется значительное различие между породами в формировании головного мозга у новорожденных и взрослых животных (Негге/ Stephan, 1955).
Значительные породные различия у собак, которые особенно проявляются в форме головы, приводят к формированию различной формы головного мозга. Длинноголовые породы имеют удлинённый рострально головной мозг, в то время как короткоголовые породы имеют более шарообразный головной мозг. Bulbus olfactorius у последних располагается под лобной долей головного мозга, в то время как у первых рострально выделяется. Gyrus proreus, соответственно, у длинноголовых пород значительно развита. Породные признаки головного мозга развиваются постнаталь-но и выражаются в основном у взрослых животных. Более подробно см. у Herre/Stephan (1955). Также и у кошек после рождения происходит изменение картины извилин (Tilney, 1934). Более подробную информацию по пре- и постнатальному развитию борозд головного мозга у кошек и собак см. у Morawski (1912).
У собак латеральная щель большого мозга, fissura lateralis cerebri, окружена двумя бороздами, эктосильвиевой, sulcus ectosylvius (rostralis et caudalis) и надсильвиевой, sulcus suprasylvius, на которой выделяют 3 части (rostralis, medius et caudalis). Выше проходит, начинаясь от каудальной поверхности полушарий, маргинальная борозда, sulcus marginalis. Рострально она переходит в выпуклую венечную борозду, sulcus coronalis, от которой в области перехода ростромедиально отделяется петлевая борозда, sulcus ansatus. В каудальной части sulcus marginalis по бокам идут две короткие борозды, sulcus ectomarginalis и (не всегда выраженная) sulcus endomarginalis.
На дорсальной поверхности полушарий имеется крестовидная борозда, sulcus cruciatus. Она проходит на медиальную поверхность и заканчивается поясной бороздой, sulcus cinguli, sulcus cruciatus аналогична sulcus centralis у людей, sulcus proreus ответвляется рострально от sulcus praesylvius.
На медиальной поверхности sulcus cinguli переходит в борозду валика, sulcus splenialis. Надвали-ковая борозда, Sulcus suprasplenialis располагается дорсокаудально. Рострально от мозолистого тела располагается борозда колена, sulcus genualis, а также sulcus ectogenualis.
По Cohn/Papez (1933) у 64 — 100 исследованных собак встречается задняя шпорная щель, fissura calcarinaposterior (caudalis), которая соответствует каудальной горизонтальной ветви sulcus splenialis.
Доместикация, в целом, приводит к уменьшению объёма головного мозга, по сравнению с дикими формами (Kruska, 1980). Однако в процессе доместикации у собак в лобной доле количество извилин увеличивается, в то время как затылочные доли сходны с таковыми у диких форм. Возраст и пол не влияет на количество извилин мозга (Klatt, 1921). По различиям картины извилин мозга у собак см. Filimonoff (1928), Marburg (1934) и Oboussier (1949).
Извилины у кошек построены проще. Sulcus ectosylvius прерывается, средняя часть не выражена. Sulcus ectomarginalis отсутствует, также как и sulcus proreus и sulcus ectogenualis. sulcus coronalis не связана с sulcus marginalis.
У кошек всегда имеется fissura lateralis cerebri и sulcus cruciatus', могут отсутствовать sulci postcruciatus, genualis et marginalis, причём это может наблюдаться, в основном, на какой-либо одной стороне (Kawamura, 1971). Sulcus diagonalis может идти горизонтально или вертикально.
Строение борозд определяет строение извилин. Извилины носят такие же названия, как и борозды, или опирается на них. Полушария также разделяются на доли, lobi. Также следует иметь в виду, что в строении борозд и извилин имеются индивидуальные вариации.
По nomina anatomica veterinaria gyrus suprasylvius отсутствует.
Gyrus suprasylvius rostralis соответствует gyrus ectomarginalis rostralis;
Gyrus suprasylvius medius — gyrus ectomarginalis medius, pars lateralis;
Gyrus suprasylvius caudalis — gyrus ectomarginalis caudalis.
Gyrus ectomarginalis называется как gyrus ectomarginalis medius, pars medialis.
Кора головного мозга, cortex cerebri, у собак имеет толщину между 1. 28 мм (затылочная доля) и 2. 25 мм (височная доля).
Neopallium обычно характеризуется наличием 6-слойной коры, cortex (neocortex, isocortex), число слоев palaeopallium и archipallium варьирует (palaeo-cortex, archicortex, allocortex).
Число и толщина слоёв, как и расположение и вид нервных клеток, меняются. В коре мозга различают поля, аrеае.
Разделение на поля и сегодня проводится по Brodmann (1909), который у людей выделяет 52 поля и обозначает их цифрами. Так, для домашних млекопитающих животных применяют отдельные обозначения, например, area 17 для зрения.
Описательное разделение коры головного мозга может быть дополнено функциональным разделением коры. По клиническим и экспериментальным исследованиям в коре головного мозга локализуются определённые функции.
Восприятие возбуждений от органов чувств осуществляется в строго определенных участках cortex cerebri. Так, органы чувств через свои рецепторы проецируются в коре головного мозга. Соответствие в деталях и локализация широко известны (например, участков сетчатки и волосковых клеток внутреннего уха). Это также касается механорецепторов кожи, что позволяет отобразить всё тело животного в коре мозга.
Отсюда следует, что поверхность полушарий находится в связи с размером тела животных (поверхность тела). Поэтому крупные животные имеют богатый извилинами мозг, в отличие от небольших животных. Также на строение коры оказывает влияние уровень организации. Так, у крупных животных с наибольшим уровнем организации мозг имеет больше извилин (напр., плотоядные — приматы).
Сведения о проекции на кору мозга настолько точные, что акустические частоты, принимаемые во-лосковыми клетками улитки, cochlea, принимают в кГц, локализуются в области 47 в полушариях.
Также имеется корреляция полей с определёнными группами мышц.
Хотя поля коры головного мозга не служат точными ориентирами границ функциональных областей коры и не соответствуют им.
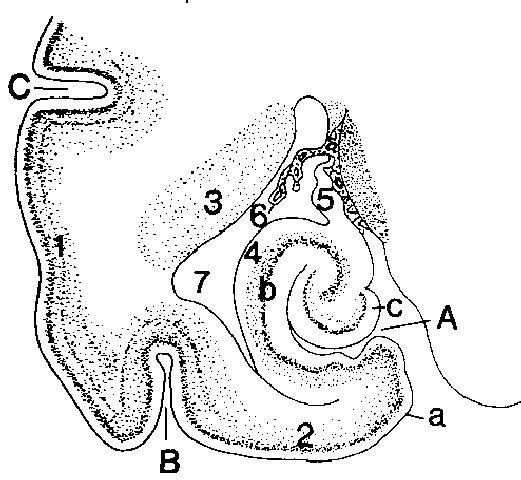
Рис. 12. Полусхематическое изображение поперечного сечения левого гиппокампа собаки (по Seiferle, 1992)
A sulcus hippocampi; В sulcus rhinalis lateralis; С sulcus suprasylvius
a gyrus parahippocampalis; b hippocampus; с gyrus dentatus
1 isocortex (новая кора); 2 allocortex (примитивная кора); 3 nucleus caudatus (хвостатое ядро); 4 alveus (лоток); 5 fimbria hippocampi (бахромка); 6 plexus chorioideus ventriculi lateralis (сосудистое сплетение бокового желудочка); 7 cornu temporale ventriculi lateralis (височный рог бокового желудочка)
Чувствительное поле коры, area sensoria, располагается в области gyrus postcrucia tusvi gyrus ectomarginalis rostralis. Оно принимает афферентные волокна от противоположной поверхности тела, во вторичное поле от обеих поверхностей тела.
Зрительное поле, area optica, у собак проходит через область gyrus splenialis и каудальную часть guri marginalis et ectomargynalis, а также gyms occipitalis.
Слуховое поле, area acustica, у собак располагается в gyrus ectosylvius и gyrus sylvius. Частоты в 100—400 Гц воспринимаются в gyrus ectosylvius rostralis, 400 — 8000 Гц — в gyrus ectosylvius medius, a 8000 — 16000 Гц — в gyrus ectosylvius caudalis.
Вкусовое поле, area gustatoria, с общей висцеральной чувствительностью локализуется в островке Рейля.
Двигательное поле коры, area motoria, у собак и кошек простирается по ростральной половине gyri postcruciatus et coronalis, а также по вентролатеральной части gyrus praecruciarus до вентральной части sulcus praesylvius. По Michaels/ Davinson (1930), area motoria у кошек составляет более 3. 7 см2 (4. 9% всей поверхности полушарий), у собак выше 14. 3 см2 (6. 75%).
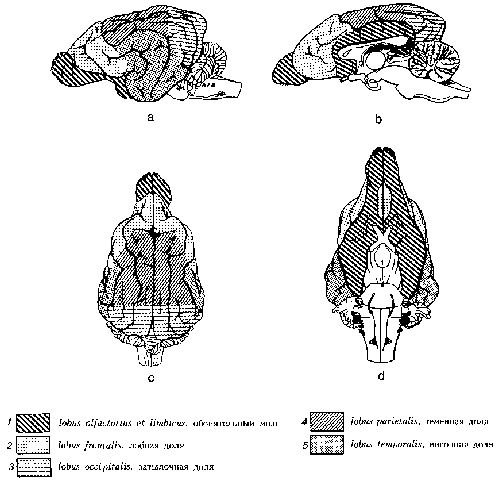
Рис. 13. Деление на доли большого мозга собак (по Seiferle, 1992)
а вид с боковой стороны слева; b разрез по срединной плоскости; с вид с дорсальной стороны; d вид с вентральной стороны
Проекционные поля дополняются ассоциативными полями, которые не имеют прямого соединения с периферией.
Лимбическая система.
Полушария окружают дугообразно двумя извилинами, gyri cinguli et parahippocampalis, мозолистое тело и формируют область разделения между neoencephalon и palaeoencephalon. Из-за этой разделительной зоны дугообразная извилина называется краевой извилиной, gyrus limbicus. Эта извилина функционально соединена с другими областями и содержит центр эмоциональных проявлений. Он контролирует психические и эмоциональные аспекты поведения и поэтому называется висцеральным, или эмоциональным головным мозгом.
Краевая система функциональная, не морфологическая, gyri parahippocampalis, cinguli et paraterminalis объединяются в lobus limbicus. К краевой системе относится area septalis, corpus amygdaloideum и гиппокамп. Между тем, эта система больше, так как также включает соединения с названными участками.
Papez впервые в 1937 году выяснил, что часть обонятельного отдела мозга отвечает не только за обоняние. Прежде всего это показывают нарушения поведения при поражении структур обонятельного мозга при бешенстве.
Гипоталамус имеет центральное место не только в контроле вегетативных функций (температуры, частоты сердцебиений, кровеносного давления, осмолярности, потребления воды и питания), но и соответствующей мотивации и стимулов, и играет большую роль в проявлении эмоций. Parez указывает на связь между гипоталамусом и cortex cerebri.
Мозговое тело, corpus medullare, состоит из переплетающихся волокон, которые могут быть частично изолированы некоторыми методами.
Ассоциативные волокна связывают области коры друг с другом.
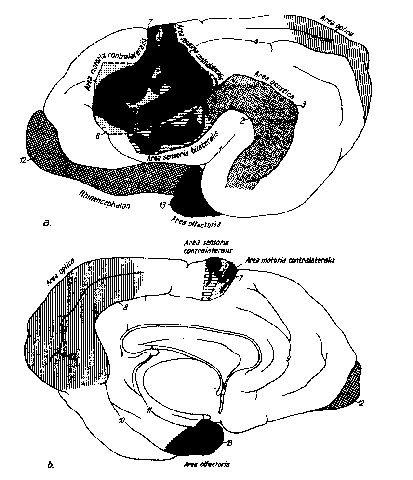
Рис. 14. Схематическое изображение двигательных, чувствительных и сенсорных областей коры головного мозга собаки, согласно экспериментальным исследованиям Campbell (1905), Marquis (1934), Pinto Hamuy et al. (1956), Tunturi (1950) и Woolsey (1960) (no Seiferle, 1992)
а вид с латеральной стороны, b вид левого полушария большого мозга с медиальной стороны 1 fissura sylvia; 2 sulcus ectosylvius; 3 sulcus suprasylvius; 4 sulcus marginalis; 5 sulcus coronalis; 6 sulcus praesylvius; 7 sulcus cruciatus; 8 sulcus splenialis; 9 sulcus genualis; 10 каудомедиальное окончание sulcus rhinalis lateralis; 10’ sulcus calcarinus; 11 sulcus hippocampi; 12 bulbus olfactorius; 13 lobus piriformis
Комиссуралъные волокна проходят поперечно и соединяют идентичные участки мозга. Ростральная спайка, commissura rostralis, — соединение palaeopallium, также внутри обонятельного мозга. Эта спайка очень развита у плотоядных животных. Спайка свода, commissura fornicis (или hippocampi), располагается между телами свод, corpora fornicis (archicorfex) в виде пластинки. Поперечной волокнистой пластинкой в neopallium являетcя мозолистое тело, corpus callosum, которое покрывает боковые желудочки, а рострально с коленом, депи, и каудально с валиком мозолистого тела, splenium corporis callosi поворачивает дугообразно к полушариям. Между мозолистым телом и столбом свода располагается прозрачная перегородка, septum pellucidum (area postcomissuralis области перегородки). Объём мозолистого тела зависит от степени развития neopallium.
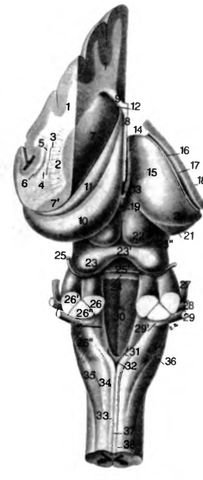
Рис. 15. Ствол мозга собаки. Вид с дорсальной стороны Слева: базальные ганглии отделены, гиппокамп сохранён, справа: виден глубоко лежащий таламус
Крыша III и IV желудочков, а также области прикрепления plexus chorioideus (taeniae chorioideae) в области боковых карманов IV желудочка на схеме не показаны
1 поверхность разреш neopallium; 2 capsula interna: 3 putamen; 4 capsula externa 5 claustrum; 6 capsula extrema; 7 nucleus caudatus, caput, 7' cauda; 8 septum pellucidum (отделена); 9 radiatio corporis callosi (отделено 4); 10 hippocampus; 11 crus fornicis; 12 ventriculus lateralis, 13 III ventriculus; 14 stria medullaris thalami; 15 thalamus; lb stria tenninalis (в основании боковых желудочков, после удаления plexus chorioideus); 17 sulcus terminalis; 18 поверхность разреза к конечному мозгу (базальные ганглии); 19 glandula pinealis; 20 corpus geniculalum laterale; 21 corpus geniculatum mediale; 22 colliculus rostralis; 23 colliculus caudalis, 23' commissura colliculi caudalis, 23" brachium colliculi caudalis; 24 velum medullare rostrale (с каудальной стороны отделено); 25 n. trochlearis, 25' decussatio n. trochlearis; 26 pedunculus cerebellaris rostralis, 26' pedunculus cerebellaris medius, 26" pedunculus cerebellaris caudalis; 27 n. trigeminus; 28 n. facialis; 29 n. veslibulocochlearis, 29' tubereulum acusticum; 30 IV ventriculus; 31 area postrema; 32 obex; 33 fasciculus gracilis; 34 fasciculus cuneatus; 35 tractus spinalis n. trigemini; 36 fibrae arcuatae superficiales; 37 sulcus medianus dorsalis (спинной мозг); 38 sulcus intermedius dorsalis (спинной мозг)
«— apertura lateralis ventriculi quarti
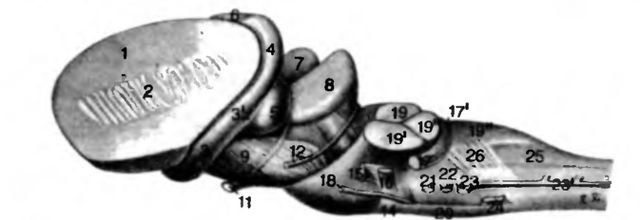
Рис. 16. Ствол мозга собаки. Вид с латеральной стороны слева
1 поверхность разреза corpus striatum; 2 capsula interna; 3 tractus opticus, radix lateralis, 3' radix medialis; 4 corpus geniculatum laterale; 5 corpus geniculatum mediale. 6 thalamus; 7 colliculus rostralis; 8 colliculus caudalis; 7+8 tectum mesencephali; 9 crus cerebri; 10 tractus cruralis transversus; 11 n. oculomotorius; 12 n. trоchlearis: 13 pons; 14 corpus trapezoideum; 15 n. trigeminus; 16 n. facialis; 17 n. vestibulo-cochlearis, 17' tubereulum acuslicum; 18 n. abducens; 19 pedunculus cerebellaris rostralis, 19' pedunculus cerebellaris medius, 19" pedunculus cerebellaris caudalis; 20 pvramis; 21 n. glossopharyngeus; 22 n. vagus; 23 n. accessorius, 23' radices spinales n. accessorii; 24 n. hyppglossus; 25 tractus spinalis n. trigemini; 26 fibrae arcuatae superficiales
Проекционные волокна сформированы в восходящие и нисходящие пути. Они достигают и покидают полушарие через внутреннюю капсулу, capsula interna, в области границы конечного и промежуточного мозга и продолжаются в ножки мозга, crus cerebri. Таким проекционным путём является tractus corticospinalis. Стволовая часть telencephalon выходит из соединения с diencephalon. Так называемые базальные ганглии лежат в глубине островка. К neencephalon относится хвостатое ядро, nucleus caudatus, а также скорлупа, putamen, к обонятельному мозгу в широком смысле слова (palaeopallium и archi pallium) — миндалевидное ядро, corpus amygdaloideum. Через ганглиевый холм проходят соединения между конечным и промежуточным мозгом. Волокна и внутренняя капсула, capsula interna, придают поверхности среза через эту область полосатую структуру, а этому участку ствола мозга название corpus striatum. Также выделяется часть таламуса, бледное ядро, pallidum (globus pallidum), приближающееся к скорлупе, putamen. Возникающее таким образом чечевицеобразное ядро, nucleus lentiformis, относится к двум различным участкам мозга.
Вне corpus striatum располагается наружная капсула, capsula externa, которая отделена полоской серого вещества, оградой, claustrum (-/5), от располагающейся под островком концевой капсулы, capsula extrema (-/6).
Хвостатое ядро, nucleus caudatus, располагается латерально от sulcus terminalis, выпячивается головкой, caput, в боковой желудочек и сопровождает своим суженным хвостом, cauda (-/7), дугообразно боковой желудочек.
Чечевицеобразное ядро, nucleus lentiformis, располагается латерально от nucleus caudatus и отграничен от capsulae interna et externa. Оно состоит из скорлупы, putamen из neencephalon и бледного ядра, pallidum из diencephalon.
Ограда, claustrum, проходит в виде тонкой пластинки серого вещества латерально от скорлупы под островком и отделяется от него capsula extrema (-/6).
Миндалевидное тело, corpus amygdaloideum, располагается базально, выпячивается в виде грушевидных долей, lobus piriformis наружу, а около височных рогов, cornu temporale — внутрь. Оно относится не топографически, а функционально к обонятельному мозгу.
Nucleus caudatus и nucleus lentiformis являются частями субкортикальной моторной системы, к которой относится подталамус, а особенно сетчатое образование (с nucleus ruber) среднего мозга. Эти центры у домашних млекопитающих представляют собой важные двигательные центры с отходящим от них, особенно у людей, кортикоспинальным путём, tractus corticospinalis (пирамидным путём), что противопоставляется высокоразвитой произвольной моторике в виде экстрапирамидально-двигательной системы.